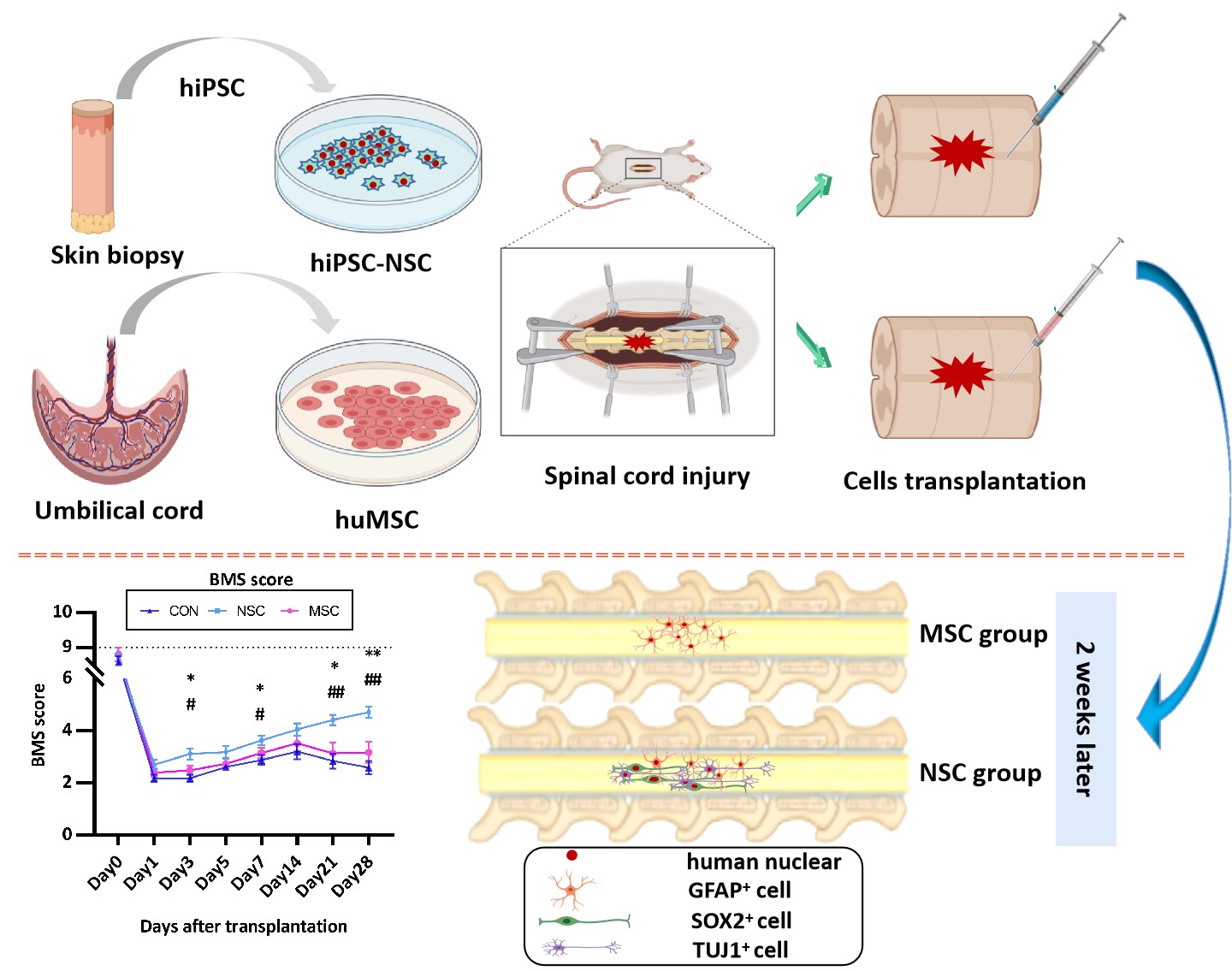
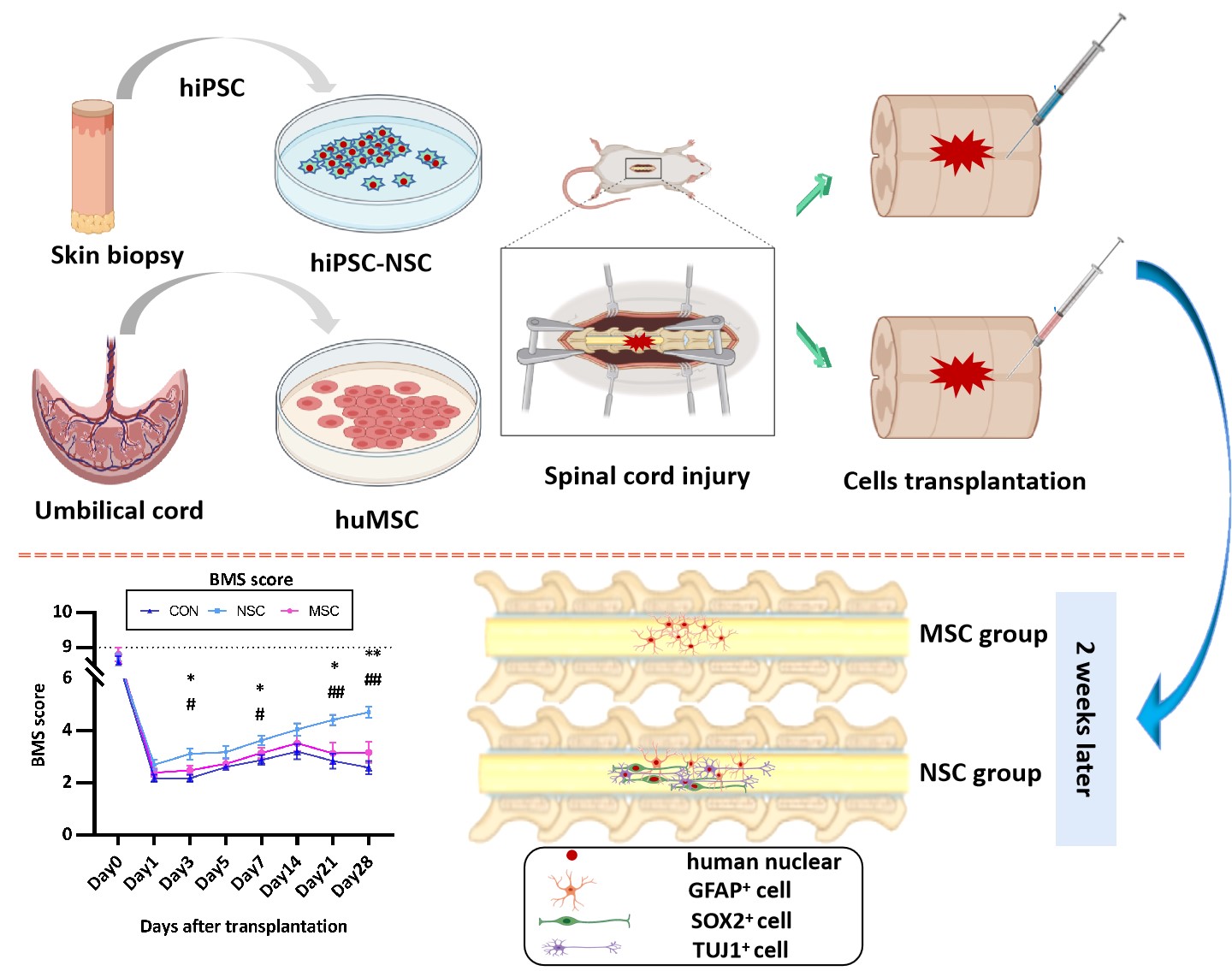
Spinal cord injury (SCI) is currently the most difficult traumatic neurological condition to treat in the clinic. Following the primary injury, which causes immediate structural damage, a series of secondary injuries, including haemorrhage, enema, demyelination, and axonal and neuronal necrosis, occur[20]. Cell transplantation is a promising treatment, including NSCs/NPCs[21], MSCs[22], and Schwann cell-like cells[23]. To date, three main sources of NSCs have been described: direct isolation from the primary CNS tissue (either from the foetal or adult brain), differentiation from pluripotent stem cells, and transdifferentiation from somatic cells. Notably, iPSC-derived NSCs have striking advantages over ESC-derived NSCs, namely, the possibility of autologous transplantation[24]. Recent investigations have also examined the utility of MSCs recovered from tissues other than the bone marrow, including the umbilical cord and adipose tissue[25]. In the present study, we transplanted hiPSC-NSCs or huMSCs into the spinal cord following SCI.
The fibrous glial scar formed by infiltrated inflammatory cells (including microglia, fibroblasts, and reactive astrocytes) limits axon regeneration across the lesion[26, 27]. The scar tissue not only exerts a protective effect by limiting the spread of inflammation and secondary damage to adjacent intact tissue but also serves as an inhibitory barrier for axon regeneration[28, 29]. Most studies conducted to date have focused on the glial scar component of the scar tissue, while the fibrotic component has received less attention. In the present study, hiPSC-NSC and huMSC transplantation reduced spinal cord fibrosis. The hiPSC-NSCs were more effective, although the difference between hiPSC-NSCs and huMSCs was not significant. Fibrotic components are not expressed at high levels in the normal adult spinal cord, and previous studies have suggested that the excess deposition observed after SCI might be attributed to various sources, such as reactive astrocytes, macrophages, and fibroblasts[28, 30]. The fibrotic changes among the different groups were similar to changes in the GFAP levels, with the lowest levels observed in the hiPSC-NSC group.
The fate of transplanted cells is determined by the environment into which the cells are transplanted rather than the intrinsic properties of the cells. In our study, allogenic MSCs survived in injured spinal cords and integrated into the host tissue without the requirement for administering immunosuppressive agents. In the present study, the transplanted huMSCs were positive for GFAP immunostaining at dpo 14, thus confirming the differentiation of huMSCs into the astrocytic lineage. The huMSCs have been confirmed to differentiate into CNS cell lineages in a rat SCI model[28, 30, 31]. Some authors investigated BM-MSC transplantation in dogs[32, 33]. Among them, Ryu et al.[32] recorded improved neurological outcomes in MSC groups after acute transplantation (one week after trauma), since all dogs exhibited purposeful hind limb motion. Moreover, some MSCs expressed markers for neurons (NF160), neuronal nuclei (NeuN) and astrocytes (GFAP)[34]. Based on our data, the transplanted hiPSC-NSCs differentiated into neuronal nuclei (NeuN)- and β-tubulin isotype III (β III tubulin)-positive neurons and glial fibrillary acidic protein (GFAP)-positive astrocytes. The most common approach for stem cell transplantation in mouse models is to inject the cells into the epicentre of the injured site. However, the tissue environment of the injured CNS is unfavourable for graft cell survival[35–37]. The number of transplanted cells gradually decreases and cells disappear from the spinal cord 3 weeks after injection[9]. When the NSCs or MSCs were transplanted in the acute phase after injury, the stem cells were no longer detected at dpo 28.
SCI induces an increase in the expression of pro-inflammatory cytokines (TNF-α, IL-6, and IL-1β), while it decreases the expression of the anti-inflammatory cytokine IL-10 [38]. As shown in the study by Cheng et al. [5], the NSC medium was able to suppress the expression of the pro-inflammatory cytokine IL-6. In the present study, NSC transplantation decreased the IL-6 level, which promoted recovery and improved locomotion. However, the level of IL-6 in the MSC group was similar to the control group. In the course of healing from the injury, the VEGF level detected in the NSC group at dpo 28 was lower than at dpo 14. A significant difference in serum TNF-α levels was not observed among the different groups at different time points. In mouse models of SCI, TNF-α-expressing cells were initially observed around the injured site at 30 to 45 min after injury, and TNF-α expression was substantially increased from 3 to 24 hrs[539, 40]. In our study, TNF-α was initially detected at dpo 7, and thus we may have missed the change in its expression. Pro-inflammatory cytokines are released from damaged cells; reactive microglia then recruit immune cells from broken blood vessels and promote tissue repair. Because reactive microglia and recruited macrophages are the main inflammatory cytokine-producing cells, the inflammatory response is then further exacerbated. The administration of IL-6, which is an important trigger of inflammatory cytokine production, to the injured spinal cord causes increased recruitment of neutrophils, an expansion of the areas occupied by macrophages and activated microglia, and reduced axonal regeneration [41, 42]. Therefore, inflammatory cytokines are strongly associated with the secondary damage after SCI.
Immunofluorescence staining showed that hNSCs and huMSCs differentiate into neurons and astrocytes; however, the functional outcome differed between the NSC and the MSC group, and the NSCs produced a better outcome than PBS and MSCs. The results are consistent with the morphological changes. A positive effect of the huMSC treatment on function was observed, but the difference was not statistically significant. The results differed from other studies[43, 44]. These opposing results are potentially attributed to the time of transplantation. Moreover, hiPSC-NSCs promoted functional recovery in SCI-induce mice not only primarily through cell differentiation and direct replacement of the lost cells but also through neuroprotective mechanisms, such as immunomodulatory factors to enhance axonal growth, modulate the environment, and reduce neuroinflammation[13].
{kind=link}
{kind=link}