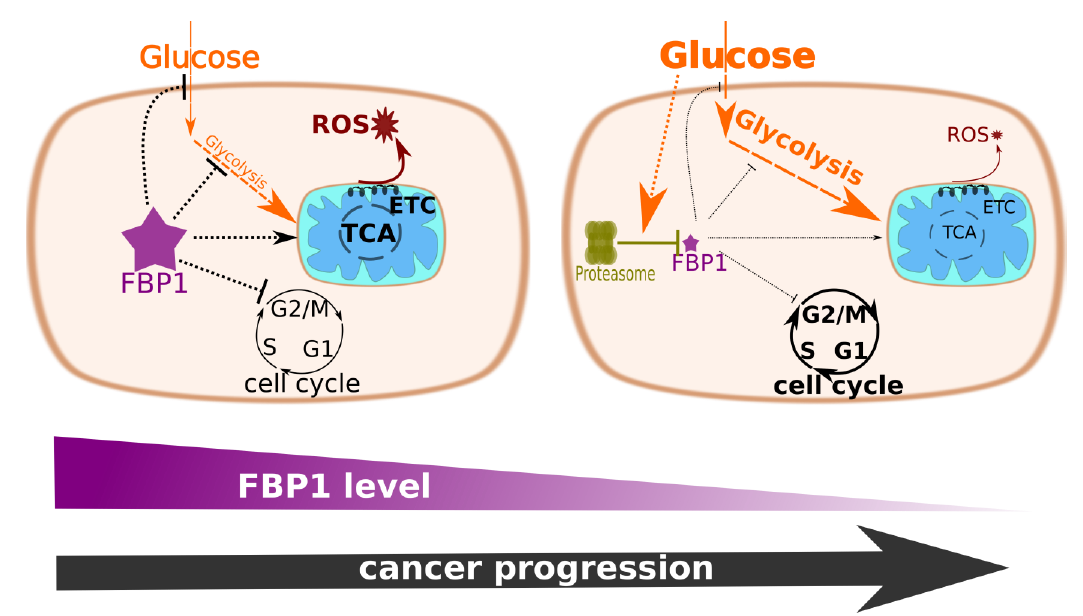
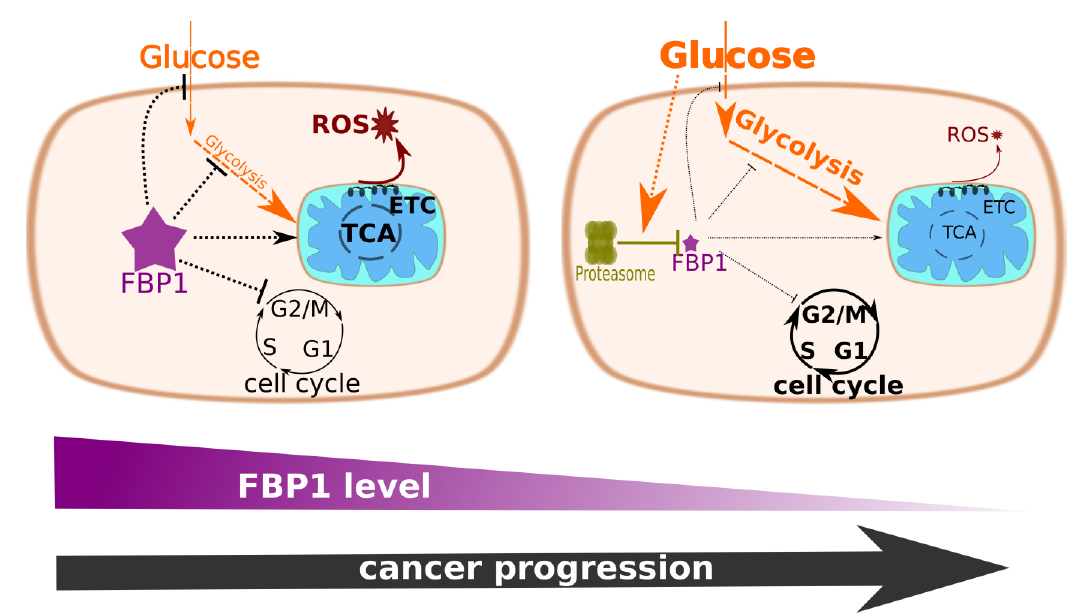
The ectopic expression of FBP1 slows down MCF-7 proliferation in presence of abundant glucose
Previous reports indicated an anti-proliferative effect of FBP1 in various cancer types [1–3, 5–7]. To assess the effect of FBP1 on proliferation, we used the protein-based SRB viability assay to assess the growth rate of stable MCF-7 lineages, in which we established various contexts of FBP1 (Fig S1). Cells with FBP1 over-expression exhibited slower growth than the vector control cells (Fig. 1a). Consistently, knocking out FBP1 increased the growth rate of MCF-7 cells with stable CRISPR-Cas9 deletion compared to the unmodified wild type cells (Fig. 1a). The changes in growth rates were not reflected in the colony formation assay (Fig. 1c). This last finding indicates that FBP1 does not hamper tumorigenesis in this luminal breast cancer subtype, consistent with the positive FBP1 expression in primary and luminal breast cancers [2].
FBP1 is essential for glucose autotrophy in MCF-7 cells
Our previous work on yeast established that FBP1 is indispensible for a continued gluconeogenesis flux [15, 16]. We therefore assessed the proliferation and survival of FBP1-KO cells under glucose depletion. Glucose withdrawal specifically impacted FBP1-KO growth and survival while the wild-type cells continued proliferating. Complete glucose deprivation inhibited the proliferation of FBP1 KO cells starting at 3 days after glucose withdrawal (Fig. 1b). Consistently, limiting glucose in medium to 2mM had comparable effects on FBP1-KO. On the contrary, neither diminished glucose nor complete glucose deprivation affected the proliferation of the wild type MCF-7 cells (Fig. 1b). This result highlights both the indispensability of FBP1 activity for gluconeogenesis in MCF-7, and also ensures that FBP1 is the only isozyme in these cells capable of catalysing the dephosphorylation of F1,6bP.
FBP1 inhibits glucose uptake under a variety of conditions in MCF-7 cells
To test the impact of FBP1 on glucose uptake in MCF-7 cells, we used 2-NBDG (fluorescence labelled 2-DG). The over-expression of FBP1 inhibited glucose uptake in full DMEM, meanwhile FBP1 KO showed a general trend towards increased glycolysis (Fig. 2a). FBP1 over-expression also inhibited glucose uptake upon glucose replenishment and under glutamine deprivation (Fig. 2b)., consistent with the reports on the anti-glycolytic effect of FBP1 in various types of cancer [2–8].
FBP1 loss increases glucose utilisation in glycolysis, pentose phosphate pathway and amino acid synthesis
In line with earlier reports on FBP1 repression as an essential step towards Warburg metabolic rewiring [2, 3, 5], we used HPLC-MS metabolite quantification to investigate the metabolic shifts in FBP1 KO MCF-7 cells in contrast with the parental wild-type cells. FBP1 KO cells demonstrated an increased abundance of glycolysis and pentose phosphate pathway intermediates (Fig. 2c,d). Increased intracellular and extracellular lactate also indicates an increased glycolytic flux. Unlike the other glycolytic intermediates, phosphoenolpyruvate and pyruvate were both depleted in the FBP1 KO cells. The increase of the intracellular and extracellular lactate on expense of pyruvate is a clear sign of increased aerobic fermentation in the absence of FBP1 (Fig. 2c,g). Since pyruvate the most net consumed metabolite by cells (Fig S2), the depletion of pyruvate in medium and the increase in intracellular and medium lactate strongly signifies an overall increase in pyruvate conversion to lactate fuelled by both glycolytic and extracellular pyruvate. Altogether this indicates glucose to lactate metabolic flux, hence a potentiated Warburg effect in the FBP1 KO cells. The depletion of pyruvate and phosphoenolpyruvate in FBP1 KO MCF-7 also coincided with an increase in alanine levels in medium, for which pyruvate is the biosynthetic precursor. Glutamine is the major nitrogen source and amino-acid in DMEM, with 2-D cultured cells showing higher reliance on glutamine compared to spheroids or in-vivo tumours [17]. The increased level of medium glutamine, observed for the MCF-7 KO cells (Fig. 2g), signifies a shift from amino acid to glucose catabolism upon FBP1 loss. Consistently, the overall output of extracellular alanine, which is net released from the cells into the medium (Fig S2), is increased, hence signifying an increased overall amino acid biosynthesis. Moreover, leucine, isoleucine, valine, tyrosine, tryptophan and phenylalanine, all essential amino acids, showed elevated accumulation in FBP1 KO cells hence indicating increased uptake and/or decreased catabolism due to the complete inhibition of gluconeogenesis (Fig S2).
A combination of GAPDH inhibition and of PKM2 activation causes serine depletion in FBP1 KO MCF-7
Intriguingly, and contrary to earlier findings [2], FBP1 KO correlated to a significant depletion of intracellular serine and its downstream derivative glycine (Fig. 2f). This depletion was not accompanied by shifts of either serine or glycine medium concentrations hence ruling out changes of uptake or secretion (Fig S2), while the activity of PHGDH, the key enzyme for serine biosynthesis [18], remained unchanged (Fig S3). Changes in glycolytic enzymatic activity offer a plausible explanation for the unexpected depletion of serine in FBP1 KO cells. We detected inhibited GAPDH activity and increased pyruvate kinase activity in the native protein extracts of the FBP1 KO cells compared to wild type (Fig S3). GAPDH inhibition is a reported mechanism of fuelling the pentose phosphate shunt with G6P [19–21], and it consists with the observed increase of upper glycolytic (Fig. 2c), and pentose phosphate pathway intermediates (Fig. 2d). FBP1 substrate F1,6bP is a robustly established allosteric activator of PKM2 [22, 23], hence the removal of FBP1 spares its substrate allowing the observed activation of PKM2. Together, inhibited GAPDH and activated PKM2 deplete the glycolytic intermediates in between the two enzyme, including 3-PG, the initial substrate of serine biosynthesis (Fig. 2c&f).
FBP1 impedes glucose sensing and uptake and in MCF-7 cells
To further assess the impact of FBP1 on glucose sensing, we probed TXNIP expression. TXNIP expression is indirectly induced by the glycolytic intermediates Glucose-6-phosphate G6P and glyceraldehyde-3phosphate GA3P [24]. Hence TXNIP is completely depleted under glucose deprivation. Under continuous glucose supply, and upon glucose replenishment, FBP1 KO cells exhibited higher levels of TXNIP compared to wild type cells (Fig. 3a,b&c). Consistently, lower levels of TXNIP expression were observed in FBP1 KO cells upon glucose replenishment with varying concentrations (Fig. 3d). To show that increased TXNIP expression is a direct outcome of increased glycolysis intermediates, 2-DG was included as a positive control and elicited sharp increase in TXNIP expression (Fig. 3d).
FBP1 correlates with increased ROS accumulation, mitochondrial biomass and decreased mitochondrial inner-membrane potential MIMP
In light of the observed effects on glucose catabolism and utilisation, we sought to assess the impact of FBP1 on the abundance and function of the mitochondria in addition to the related outcome of ROS accumulation. FACS analysis of DHE stained MCF-7 cells showed increased ROS accumulation in oeFBP1 vs vector control cells; consistently FBP1 KO cells exhibited significantly decreased ROS accumulation (Fig. 4a). Using MitoTracker® Green we similarly established a positive correlation between FBP1 expression and mitochondrial biomass (Fig. 4b), indicative of increased mitochondrial biogenesis in presence of FBP1. However, the increase in mitochondrial biomass with FBP1 expression also coincided with a diminished mitochondrial inner-membrane potential MIMP (Fig. 4c).
In line with the metabolic shift towards increased glucose catabolism and decreased glutamine consumption, we predicted that FBP1 loss would render cells independent of glutamine and more vulnerable to glucose starvation. Indeed, FBP1 absence prevented the additional ROS induction observed in glutamine deprived wild type cells (Fig. 4d), meanwhile the FBP1 KO cells responded to glucose withdrawal by dramatically increasing their mitochondrial biomass to levels significantly higher than the wild type (Fig. 4e). Therefore, suggesting a compensatory response to cope with glucose scarcity by increasing mitochondrial biomass to harness higher levels of energy form a dwindling level of metabolic intermediates.
The increase in mitochondria with lower MIMP can explain the elevated ROS as result of a concomitant increase in the overall mitochondrial inner-membrane area, as a result of increased mitochondrial biomass, and leakiness in the electron transport chain. This also consists with the FBP1 inhibitory effects on mitophagy, since electron transport chain leaking is a hallmark of aging mitochondria.
FBP1 deletion diverts ATP production from the ETC
Blocking ATP-synthase with 10µM oligomycin [25] resulted in significantly smaller increase of mitochondrial polarisation in FBP1 KO MCF-7 cells compared to the wild type (Fig. 4f). The increased polarisation reflects directly the amount of protons that would path through ATP-synthase and contribute to ATP production; hence we conclude that FBP1-deficinecy limited the reliance of MCF-7 on the electron transport chain ETC for ATP production.
This consists with the increased glucose consumption, utilisation and aerobic fermentation observed in the FBP1-KO cells.
FBP1-loss provides a survival advantage under extreme hypoxia
The increased glucose utilisation and aerobic fermentation we observed for FBP1 KO cells consist with the conventional Warburg effect, and should coincide with decreased reliance on respiration. Hence we investigated the survival of MCF-7 FBP1-KO cells under hypoxia compared to wild type cells. Both SRB growth/survival assay (Fig. 5a&b) and FACS analysis of PI-stained cells (Fig. 5c) showed a statistically valid advantage for FBP1-deficeint MCF-7 cells over the wild type cells following incubations at 1% O2 saturation, SRB assays were for a time frame of 24h to 96h under hypoxia, while PI staining was performed following 48h under 1% O2 saturation.
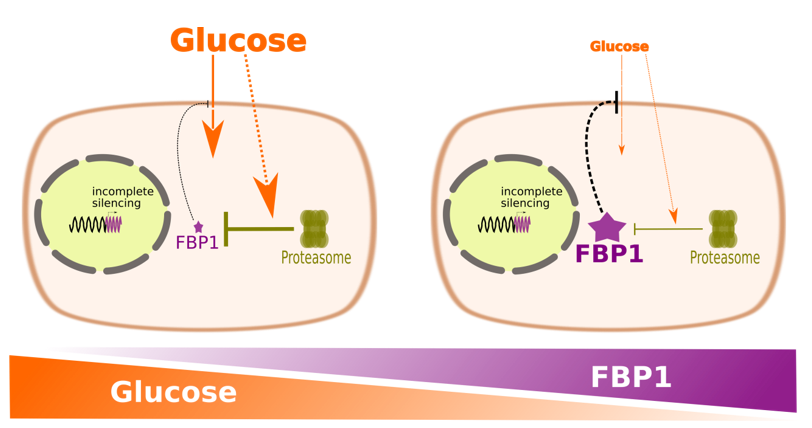
MDA-MB-231 cells acquire the capacity to degrade ectopic FBP1 in a proteasome dependent mechanism upon long-term selection
To better understand the metabolic and proliferative advantages of FBP1 silencing, we aimed to determine the effects of ectopic FBP1 in the triple-negative MDA-MB-231 cells, with depleted FBP1 expression. To our surprise the stable expression of FBP1, following a minimum of 6 weeks of geneticin selection, yielded diminished levels of FBP1 expression compared to the FBP1-positive MCF-7 (Fig. 6a). This was despite transient expression in MDA-MB-231 successfully yielding substantial levels of expression (Fig. 6b). Quantitative PCR also showed an increase in FBP1 mRNA levels in MDA-MB-231 stable oeFBP1 cells, greater than that observed in MCF-7 oeFBP1 (Fig. 6c).
Altogether this indicated the potential of post-translational protein degradation as an underlying mechanism of the attenuated FBP1 expression in MDA-MB-231 cells.
To test this possibility we resorted to pharmacological proteasome inhibition [26]. Indeed, bortezomib treatment rescued FBP1 protein expression in MDA-MB-231 cells with stable FBP1 ectopic expression while showing no effect on FBP1 in the control MDA-MB-231 cells harbouring the empty vector (Fig. 6d). To verify the rescuing effect of proteasome inhibition on ectopic FBP1 expression in MDA-MB-231 cells we also used MG132, another more distinctive proteasome inhibitor [27], with comparable outcome to that observed with bortezomib (Fig. 6e). We are the first to observe such degradation of FBP1 in breast cancer cells, a similar effect has been found to contribute to FBP1 ablation in hepatocellular carcinoma [28].
Upon bortezomib treatment, the rescued ectopic FBP1 expression also inhibited glucose uptake compared to the vector control cells (Fig. 6f), similar to its identified role in MCF-7. Moreover, the transient transfection with FBP1, correlated to higher levels of expression, decreased acidification rates in MDA-MB-231 cells indicting slowed down glycolysis (Fig S4).
Glutamine as an exclusive carbon source stabilizes ectopic FBP1 expression in MDA-MB-231 cells and confreres a growth disadvantage to oeFBP1 cells
We next sought to assess the interplay between the ectopic FBP1 expression in MDA-MB-231 and nutrient availability in the culture medium. The post-translational proteasome-dependent degradation of FBP1 has long been described as a part of the more encompassing phenomenon of carbon catabolite inactivation in unicellular eukaryotes [29]. Glucose is the most prominent metabolite to trigger the signalling cascade, which leads to FBP1 phosphorylation, ubiquitination, and degradation both in the proteasome and the vacuole [30, 31]. Therefore, we asked whether this mechanism is conserved in human cells. If so, the constitution and balance of carbon sources in cell culture medium should influence the observed levels of ectopic FBP1 by altering its degradation rate.
First, we observed via proliferation SRB assays that the marginal amounts of stably expressed FBP1 in MDA-MB-231 were advantageous under full DMEM containing both glucose and glutamine (Fig. 7a), as well as under high glucose concentration (75mM) with NH4Cl as the exclusive nitrogen source (Fig. 7b), the successful utilisation of ammonia by breast cancer cells has been clearly demonstrated [32]. To our surprise however, the growth of oeFBP1 MDA-MB-231 cells was severely challenged in medium with high levels of glutamine (8mM) as the exclusive carbon and nitrogen source (Fig. 7c). This is very intriguing in light of glutamine being a major gluconeogenic substrate. Consistently, immunoblotting showed remarkable increase in ectopic FBP1 levels under glucose depletion coupled with high glutamine concentrations (Fig. 7d), thus indicating an attenuation of FBP1 degradation under this condition.
This is to our knowledge the first reported evidence on the effect of the availability of various carbon sources on FBP1 proteasomal degradation in human cancerous cells and mammalian cells in general. Hence indicating a likely carbon catabolite inactivation phenomenon akin to that characterised in yeast.
Despite its advantageous effect on proliferation, the presence of marginal ectopic FBP1 in oeFBP1 cells tangibly inhibited colony formation capacity (Fig. 7e), consistent with the known aspects of FBP1 repression in basal-like and metastasising breast cancer.
Stable and transient FBP1 expression lead to contrasting oxidative and mitochondrial outcomes in MDA-MB-231 cells
Compared to its effect in MCF-7, the stable expression of FBP1 in MDA-MB-231 unexpectedly correlated to a steep decrease in ROS levels as measured using DHE (Fig S5a). However, transient expression yielding comparable levels of FBP1 to those of MCF-7 had no significant effect on ROS accumulation (Fig S5d), despite a slight trend towards increased ROS. The investigated mitochondrial markers, on the other hand, reacted similarly to the expression of FBP1 in MDA-MB-231 in a manner comparable to the observations in MCF-7. MDA-MB-231 cells with stable FBP1 expression exhibited lower mitochondrial inner-membrane polarisation and higher total mitochondrial biomass (Fig S5b&c).
Transient FBP1 over-expression however yielded elevated mitochondrial polarisation (Fig S5e), hence possibly indicating an increase in respiratory activity upon FBP1 ectopic expression later followed by increased mitochondrial biogenesis.
Comparing transcriptional profiles signified a wide-scale cell-cycle inhibitory effect of long term FBP1 expression in MDA-MB-231 cells
Having observed that MDA-MB-231 cells adapt to FBP1 long-term expression by degrading FBP1 in the proteasome, we were intrigued to assess the shifts in transcription that can indicate the types of alterations occurring as a result of ectopically expressing FBP1 in MDA-MB-231 cells. Therefore, we used the Illumina RNA microarray system for a full transcriptome scale quantification of gene-expression of stable oeFBP1 vs vector control MDA-MB-231 cells.
Following the basic quantification of fold changes in FBP1 over-expression vs vector control cells, we performed a wide analysis of the impact of FBP1 over-expression on gene-expression patterns in MDA-MB-231 including a larger pool of up-regulated (≥1.6 folds) and down-regulated (≤0.65 folds) genes (Figure S6). Using the STRING functional protein association network, two separate schemes of interaction networks between the proteins encoded by the up-regulated and down-regulated genes were made. Comparing the two interaction networks, the most noticeable difference is the clear arrangement of the down-regulated genes into an interaction-rich network (Fig S6a). At the centre of this network are proteins associated with the cell cycle progression; the vast majority are proteins essential for mitosis and cytokinesis (M-Phase) these include the centrosome proteins CENP- A,E,M,N, aurora kinase A AURK-A and its activator TPX, the kinesin family members KIFs 2C, 4A, 20A, 20B, 23 and the centrosomal protein CEP55. Besides M-phase proteins, several other genes encoding for essential cell-cycle proteins are also seen within the aforementioned interaction network. These include, the Cyclin A2 essential for the cell cycle progression G1/S and G2/M transitions through activating CDK1 (also among the down-regulated genes) and CDK2. Other down-regulated genes include the histone cluster 2 (H2AC and H2AA3) in addition to thymidilate synthetase essential for thymidine de-novo biosynthesis and the exonuclease 1 EXO1 essential for DNA-mismatch repair. In contrast with the heavy interactions observed between the down-regulated genes, the interactions among the up-regulated genes are scarce. Therefore no wider conclusions regarding the up-regulation of entire signalling pathways or cellular functions could be drawn.
Immunoblotting confirmed the FBP1-associated inhibition of CENP-E
In order to test the observed cell-cycle inhibition on a protein level, immunoblots of short-term transfected MDA-MB-231 cells were performed. HT-29 cells were also included to test the validity of our findings in other FBP1-deficeint cell lines of other tumor origins. Consistent with the transcriptional findings, the ectopic expression of FBP1 correlated to diminished CENP-E levels in both tested cell lines (Fig. 8). This finding validates the functional relevance of the cell cycle inhibition seen in the transcriptional profiling.
{kind=link}
{kind=link}
{kind=link}