Bacteriophages (phages), first used as antibacterial agents more than a century ago1, offer new hope for highly resistant infections2,3 but clinicians remain uncertain about optimal dosing and treatment duration. Here we describe phage, bacterial and host responses to intravenous (IV) phage therapy for a chronic osteoarticular infection complicating K-wire insertion for traumatic fracture-dislocation in a 7-year-old girl. Previous surgical debridement and hardware removal and three months of intravenous antibiotics had failed to stop the progress of disease (Fig. 1) and no other medical or surgical options were available. Under informed consent, we administered 1011 plaque forming units (PFU)/mL of Pa14NPΦPASA16 (PASA16) once (days 1, 2, 4–7) or twice daily (days 3, 8–14; Fig. 2a) for two weeks, beginning three months into a twelve-month course of IV colistin and aztreonam. Strong lytic activity of PASA16 against a surgically obtained isolate of P. aeruginosa (Ppa2.1; efficiency of plating [EOP] ~ 100%) was unaffected by these antibiotics in vitro (Supplementary Fig. 1).
Transiently increased heel pain 8 hours after the first dose of PASA16 was followed by fever to 39.7oC several hours later. Low-grade temperatures (37.7-37.8oC) were also recorded ~ 6–12 hours after the second and third doses of phage but none thereafter. C-reactive protein (CRP) peaked on day 4 (20.1 mg/L) (Fig. 2a) and a slight increase in serum Ig-G levels from baseline to day 30 (14.7 g/L to 16.1 g/L, normal range 6.24–14.4 g/L) was noted, with no change in serum complement levels (C3/C4). Pain-free weight bearing on the affected leg was achieved for the first time since the initial injury, seven weeks after completion of phage therapy, and long-term follow-up demonstrated clear radiological improvement (Fig. 1).
Bacterial and phage genomes were quantified in DNA extracted from whole blood and serum respectively, as previously described3 (Supplementary Text). Standard curves were generated by spiking tenfold serial dilutions of log-phase P. aeruginosa and PASA16 into whole human blood (P. aeruginosa, 1.3–1.3 × 106 colony forming units [CFU]/mL) or serum (phage, 1 × 101–7 PFU/mL). Viable phage was recovered from serum onto a lawn of an aztreonam- and colistin-resistant strain of P. aeruginosa (JIP697; EOP ~ 100%), also as previously described3. All experiments were performed in duplicate.
Blood cultures yielded no bacteria at any stage, but viable phage was recovered from pre-dose samples up until day 5 consistent with productive infection of target bacteria throughout the dosing interval (Fig. 2a). Increased phage dosing in the second week was associated with further bacterial lysis but the dynamics of bacterial and phage DNAemia (to ~ 105 genome equivalents/mL), similar to previous observations3,4 are most consistent overall with a classic “predator-prey” relationship (Fig. 2a), with predators (phage) thriving while prey (bacterial hosts) are plentiful but declining as prey populations are consumed.
Endotoxin levels in the administered phage dose were below the accepted human pyrogenic threshold of 5 EU/kg (see Supplementary Text for calculations). The expected amount of bacterial endotoxin accompanying the surges of Pseudomonas DNA (102-103/mL) into the bloodstream is orders of magnitude higher at between 0.05 and 0.5 ng/mL 5,6 (~ 40–400 EU/kg) and expected to produce a strong inflammatory response. Significant differences in the expression of 18 genes associated with the innate immune response (Gene Ontology: 0045087, p = 1.03 × 10− 11) between D0 (immediately prior to phage therapy) and D2-4 (24–72 hours after initiation of phage therapy) were demonstrated by principal component analysis (Supplementary Fig. 2; p < 0.05) and coincided with the fever, localised pain and spike in Pseudomonas DNA and CRP (Fig. 2a). Hierarchical clustering analysis (Supplementary Text) confirmed enrichment of human innate immune response-associated genes on D2 vs. D0 (Fig. 2b). The relative enrichment of seven of these (ATG5, CD180, CD244, PEZ1, IL5RA, PTGDR2, SMPD3) persisted for at least two weeks after completing phage therapy (D29), but were no longer significant beyond D5-7 after post-hoc adjustment (Benjamini-Hochberg). Gene Set Variation Analysis (Supplementary Text) further demonstrated early immediate upregulation of genes enriched for innate immune response in periods 1 (D2, D4) and 3 (D9, D11) in synch with the kinetics of Pseudomonas DNAemia, followed by upregulation in genes enriched for adaptive immune response from D5 onwards (Fig. 2c), reflected at the systems level by the hierarchical clustering analysis.
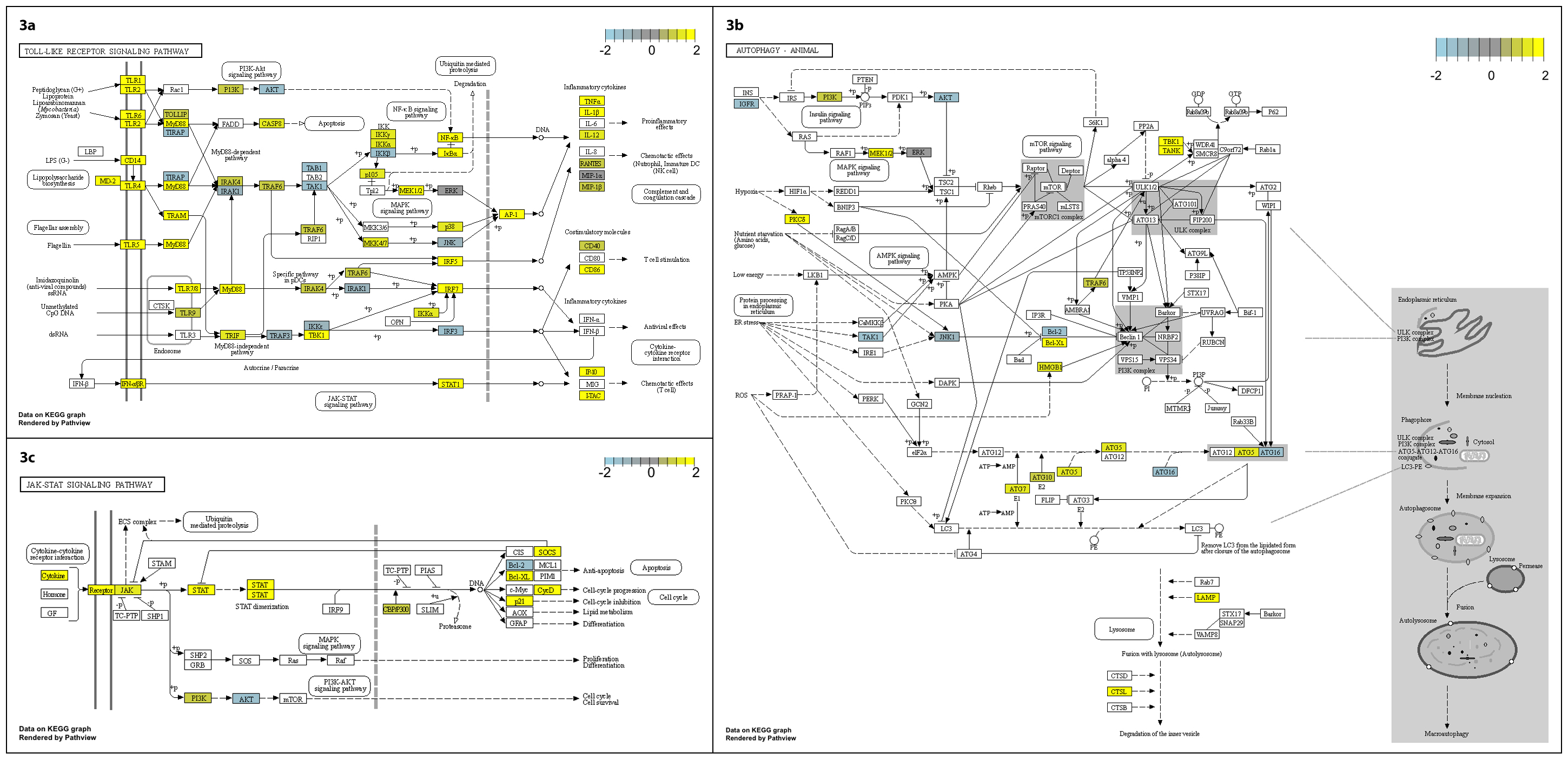
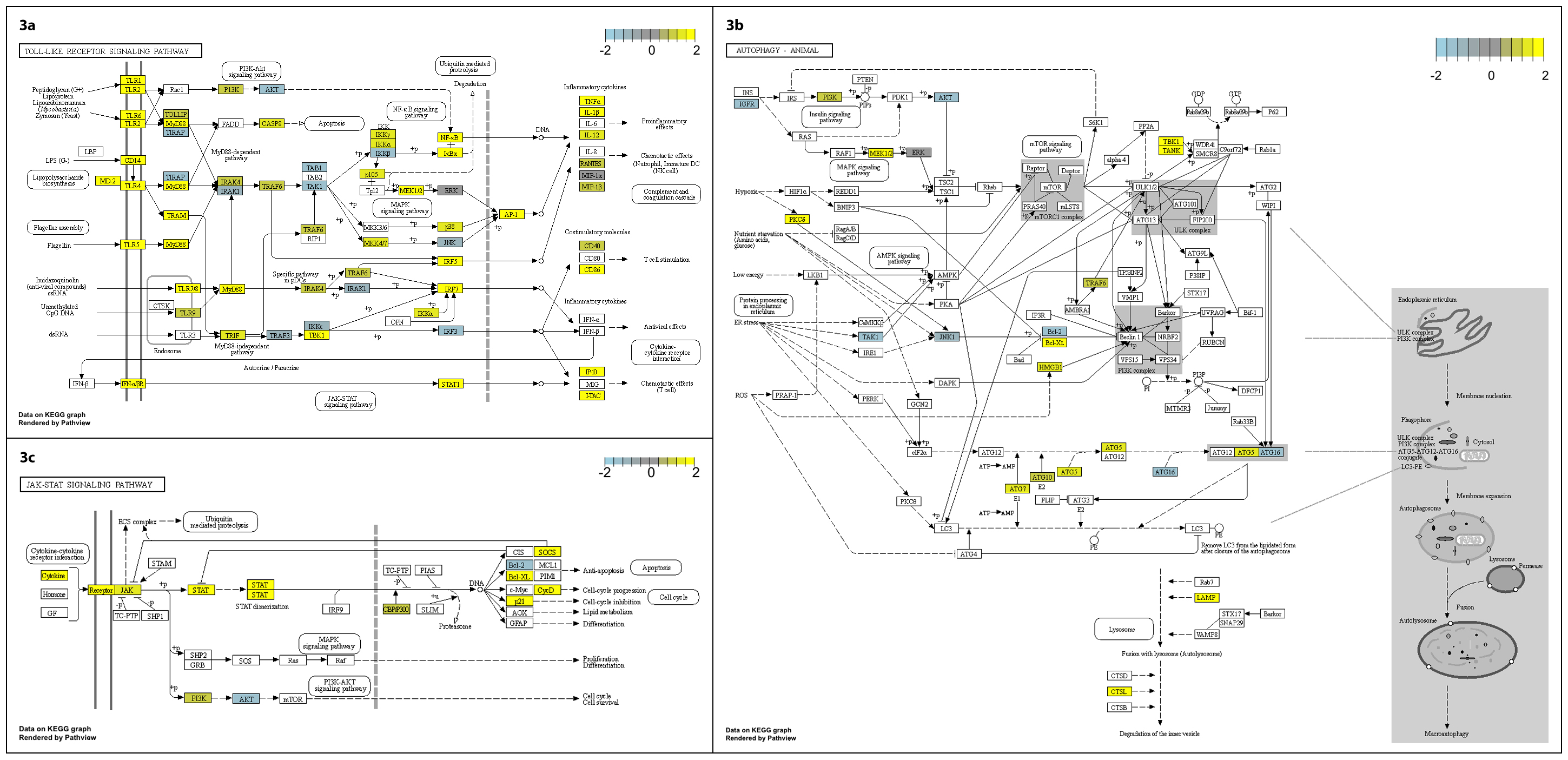
We note the differential expression of genes linked to autophagy such as ATG5, that may be beneficial in clearance of P. aeruginosa infection7, and others (CD180 and CD244), which may dampen the inflammatory response8. The kinetics of significant changes in toll-like receptor signalling, autophagy and JAK-STAT signalling pathways (Supplementary Fig. 3a-c) align temporally with the clinical signs and symptoms and phage-bacterial kinetics described above. Taken together, these data indicate an innate immune response to phage-mediated bacterial lysis. It appears that clinical signs and widely available measures such as C-reactive protein, together with simple quantitation of phage and bacterial kinetics, can be used to guide the therapeutic interaction of the three main actors: phage, bacteria, and human host. The kinetics of the response are similar to previous descriptions and indicate that most benefit accrues within the first ten days of treatment. Such multi-modal therapeutic phage monitoring is important to help us understand this intervention better.
{kind=link}
{kind=link}