The nematode Laxus oneistus did not enter suspended animation upon 24 h anoxia
To survive anoxia, nematodes enter suspended animation to suppress metabolism and conserve energy. The most notorious sign of suspended animation is the arrest of motility (Nystul et al., 2003; Chan et al., 2010; Kitazume et al., 2018).
Surprisingly, although the whole population of four tested nematode species, including C. elegans, was reported to be in suspended animation upon 10 h in anoxia (Kitazume et al., 2018), L. oneistus kept moving not only after 24-h-long incubations, but also upon 6-day-long incubations in anoxic seawater (three batches of 50 worms were incubated under each condition). Additionally, the symbiotic nematodes appeared morphologically normal (Supplemental movies 1-4).
The fact that we could not observed suspended animation, led us to hypothesize that L. oneistus evolved different strategies to survive oxygen deprivation.
Stable transcriptional profile under hypoxic or anoxic sulfidic conditions
To understand the molecular mechanisms underlying L. oneistus response to oxygen, we subjected it to various oxygen concentrations. Namely, nematode batches were incubated under either normoxic (100% air saturation; O), hypoxic (30% air saturation; H) or anoxic (0% air saturation; A) conditions for 24 h. Additionally, given that L. oneistus thrives in reduced sand containing up to 25 µM sulfide (Ott and Novak., 1989; Paredes et al., 2021), we also incubated it in anoxic seawater supplemented with < 25 µM sulfide (anoxic sulfidic condition; AS).
While transcriptional differences of its symbiont (Candidatus Thiosymbion oneisti), incubated under normoxic (O) and hypoxic (H) conditions were negligible (Paredes et al., 2021), the expression profiles of nematode batches incubated under O conditions varied so much that they did not cluster (Figure S1). Consequently, there was no detectable differential expression between the transcriptomes of O nematodes and any of the other transcriptomes (H, A or AS; Figure S1B, C). We attribute the erratic transcriptional response of L. oneistus to normoxia to the fact that this concentration is not typically experienced by L. oneistus (Ott et al., 1989; Paredes et al., 2021).
As for the expression profiles of nematodes subjected to the H, A or AS conditions, replicates of each condition behaved more congruently (Figure S1B). While we did not find any significant difference between the A and AS nematodes, only 0.05% of the genes (8 genes; Data S1) were differentially expressed between the H and A nematodes and there was no significant difference between the H and A proteomes (t-test, FDR, Benjamini-Hochberg correction, p < 0.05; Figure S2A, Data S1). However, 4.8% of the expressed genes (787 out of 16,526) were differentially expressed between H and AS nematodes, with 434 upregulated under AS and 353 genes upregulated under H conditions (Figure S1C, Data S1).
Collectively, our data suggests that L. oneistus may be ill-equipped to handle normoxic sediment, but it maintains a largely stable physiological profile under both hypoxic and anoxic sulfidic conditions. Before discussing the subset of biological processes differentially upregulated in AS versus H nematodes and vice versa, we will present the physiological processes the worm appears to mostly engage with, irrespectively of the environmental conditions we experimentally subjected it to.
Top-expressed transcripts under all tested conditions
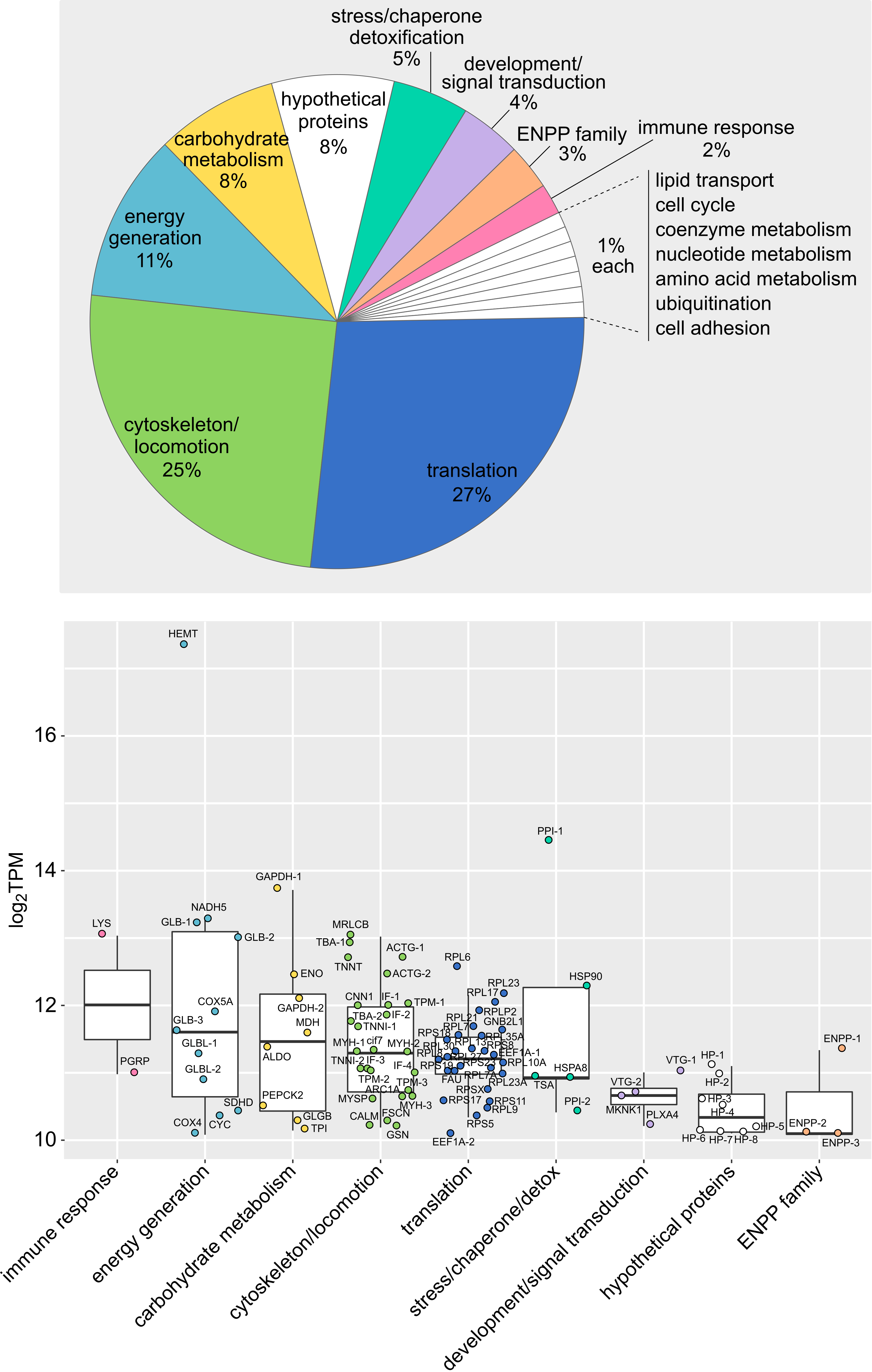
To gain insights on L. oneistus basal physiology, we treated all the 16 transcriptomes as biological replicates (i.e., O, H, A and AS transcriptomes were pooled) and identified the 100 most abundant transcripts out of 16,526 based on functional categories extracted from the UniProt database (2021) and comprehensive literature search (Figure 1, Data S2). Our manual classification was supported by automatic eggNOG classification (Data S2). Similarly, the H and A proteomes were pooled, and the 100 most abundant proteins out of 2,626 were detected (Figure S2).
Based on median gene expression values of the top 100 expressed genes, we found that some of the processes L. oneistus mostly engages with were ubiquitination (ubq-1, Stringham et al., 1992), energy generation (globin glb-1-like (Geuens et al., 2010), cytochrome c oxidase I subunit ctc-1 (UniProtKB P24893), nduo-4-like (UniProtKB P24892), stress response and detoxification (e.g., hsp-1, hsp-90, hsp12.2, and catalases ctl-1 and ctl-2; Birnby et al., 2000; Chávez et al., 2007), and immune defense (lysozyme-like proteins and lec-3) (Figure 1, Data S2).
Lastly, 48 out of the top 100 most expressed genes, were also detected among the top 100 proteins (Figure 1, Figure S2, and Data S2, Supplemental material). Despite the modest correlation between transcript and protein expression levels (r = 0.4) (Figure S3), there was an overlap in the detected biological processes (e.g., energy generation, stress response or detoxification categories, carbohydrate metabolism, cytoskeleton, locomotion, nervous system) (Figure S2).
All in all, except for those encoding for immune effectors, top-transcribed L. oneistus genes could not be ascribed to its symbiotic lifestyle. This differs to what observed for other chemosynthetic hosts, such as giant tubeworms and clams. Indeed, it is perhaps because these animals acquire their symbionts horizontally and feed on them as they are housed in their cells (and not on their surface) that they were found to abundantly express genes involved in symbiont acquisition, proliferation control and digestion (Sun et al., 2017, Hinzke et al., 2019; Yuen et al., 2019). Notably, we did observe a partial overlap of the most expressed gene categories (e.g., oxidative stress, energy generation, immune response), when L. oneistus was compared to the marine gutless annelid Olavius algarvensis. We ascribe the overlap to the fact that, albeit endosymbiotic, O. algarvensis also inhabits shallow water sand (Figure S4, Supplemental material) and, as hypothesized for L. oneistus, it may also acquire its symbionts vertically (Woyke et al., 2006; Dubilier et al., 2008; Wippler et al., 2016; Zimmermann et al., 2016).
To conclude, although both symbiont- (Paredes et al., 2021) and host-transcriptomics do not suggest a high degree of inter-partner metabolic dependence in the L. oneistus ectosymbiosis, the nematode seems well-adapted to both anoxic sulfidic (AS) and hypoxic (H) sand (Figure 2, Data S1). The transcriptional response of the worm to these two conditions is, however, significant (Figure 2, Data S1), and it will be reported next.
Genes upregulated in anoxic sulfidic (AS) nematodes
Chaperones and detoxification. The expression of chaperone-encoding (e.g., hsp12.2, grpE, dnaJ/dnj-2, pfd-1, pfd-6; Naylor et al., 1996; Lundin et al., 2008; Bar-Lavan et al., 2016), and ROS-detoxifying-related genes (e.g., superoxide dismutase sod-2 and a putative glutathione peroxidase, involved in the detoxification of superoxide dismutase and hydrogen peroxide, respectively; Suzuki et al., 1996; Margis et al., 2008) were higher in AS nematodes (Figures 2 and 3). Notably, transcripts encoding for the heme-binding cytochrome P450 cyp-13B1 were also more abundant in AS (Figure 3), perhaps to increase the worm’s capacity to cope with putative ROS formation (Oliveira et al., 2009). Indeed, as cells start being oxygen-depleted, mitochondrial ROS accumulate because of the inefficient transfer of electrons to molecular oxygen (Semenza, 1999; Nystul and Roth, 2004; Selivanov et al., 2009; Kim and Jin, 2015). Alternatively, the upregulation of antioxidant-related genes in AS worms could represent an anticipation response to an imminent reoxygenation. In animals alternating between anoxic and oxygenated habitats, the re-exposure to oxygen can be very dangerous, as it creates a sudden ROS overproduction that may overwhelm the organism’s oxidative defense mechanisms (Hermes-Lima and Zenteno-Savin, 2002; Hashimoto et al., 2004). Although it has not been reported for nematodes, overexpression of ROS-counteracting genes is consistent with what has been reported for vertebrates and marine gastropods which, just like L. oneistus, alternate between oxygen-depletion and reoxygenation (Hermes-Lima and Zenteno-Savin, 2002).
Mitochondrial and cytoplasmic ribosome biogenesis. In the cellular stress imposed by oxygen deprivation, mitochondria are central to both death and survival (Borutaite et al., 1995; Brookes et al., 2004; Brenner et al., 2012; Hawrysh et al., 2013; Galli et al., 2014). In this scenario, calcium regulation, the scavenging of ROS or the suppression of their production, and/or inhibition of the mitochondrial permeability transition pore (MPTP) opening, might help to preserve mitochondrial function and integrity (Horwitz et al., 1994; Murphy et al., 2008; Galli et al., 2014; Fanter et al., 2020). In addition, removal of specific mitochondrial components (mitochondrial-associated protein degradation, MAD), might also arise to maintain the overall mitochondrial homeostasis (Chatenay-Lapointe and Shadel, 2010; Heo et al., 2010). Perhaps as a response to anoxia-induced stress (reviewed in Galli et al., 2014), a gene involved in MAD (vms-1) (Chatenay-Lapointe and Shadel, 2010; Heo et al., 2010), was upregulated in AS worms (Figure 4). More abundant in this condition were also transcripts encoding for mitochondrial transmembrane transporters tin-44, slc-25A26 and C16C10.1 (UniProtKB O02161, Q18934, Q09461), putatively transporting, peptide-containing proteins from the inner membrane into the mitochondrial matrix, such as S-Adenosyl Methionine (Figure 6). Surprisingly, although the translation elongation factor eef-1A.2 (Tullet, 2015) was downregulated in AS worms, not only various mitochondrial ribosome structural components (28S: mrps, 39S: mrpl; Kaushal et al., 2014), and mitochondrial translation-related genes (e.g., C24D10.6 and W03F8.3; Sharika et al., 2018) were upregulated in AS nematodes, but also several cytoplasmic ribosome biogenesis (40S: rps, 60S: rpl; Melnikov et al., 2012) and subunit assembly genes (e.g., RRP7A−like, You et al., 2015) (Figure 4).
Taken together, the maintenance of mitochondrial homeostasis, an anticipatory response to a potential upcoming ROS insult (see Chaperones and detoxification section) and/or their involvement in extra-ribosomal functions (Chen et al., 2010; Savada et al., 2014; Xu et al., 2016) might explain the upregulation of ribosomal biogenesis-related genes in AS nematodes. Although upregulation of ribosomal proteins has also been observed in anoxic gastropods (Larade et al., 2001), increased ribosomal biogenesis (which oftentimes directly correlates with an increase of protein synthesis) is not expected in animals that must repress their metabolism to cope with oxygen deprivation (Thomas et al., 2000; Hochachka and Lutz 2001; Shukla et al., 2012).
Energy generation. Equally surprising was the upregulation of all differentially expressed genes related to energy generation in AS nematodes (Figure 4). Namely, besides putative oxygen-binding globulin-like genes (e.g., glb-1, glb-14, Geuens et al., 2010), the following were upregulated in AS nematodes: key structural genes (e.g., atp-3, atp-5, Xu et al., 2018), assembly-related genes (H+-transport ATP synthase, Maglioni et al., 2016) of the mitochondrial ATP synthase (complex V), genes related to complex I (lpd-5, nuo-2, McKay et al., 2003; Rea et al., 2007), a subunit of the succinate dehydrogenase involved in complexII (mev-1, Hartman et al., 2001), a mitochondrial cytochrome C oxidase subunit II assembly gene related to complex IV (sco-1, Williams et al., 2005), and a mitochondrial gene (coq-5), involved in the synthesis of either ubiquinone (Q, aerobic) or rhodoquinone (RQ, anaerobic) electron carriers (Buceta et al., 2019) (Figure 4). This suggests that, under anoxia, the electron transfer chain (ETC) is rewired in such way that electrons still enter the ETC at complex I, but instead of reaching complex III and IV they are transferred to RQ. This, in turn, shuttles the electrons to succinate dehydrogenase. The latter enzyme uses fumarate as an alternative electron acceptor, reducing it to succinate. This mechanism would maintain the flow of electrons through the ETC, and, it would prevent mitochondrial ATP generation (complex V) from shutting down (Buceta et al., 2019; Del Borrello et al., 2019).
In short, under AS, similarly to what has been observed in other free-living and parasitic nematodes, complex I appears to be the sole proton pump in this truncated form of ETC (Buceta et al., 2019; Del Borrello et al., 2019). In accordance with this hypothesis, tryptophan (Trp) degradation-related genes (acsd-1, acsd-2) and the Trp RNA ligase (wars-1; Tsai et al., 2017) that might be required to synthesize RQ (Buceta et al., 2019; Del Borrello et al., 2017; Tan et al., 2020) were upregulated under AS. Intriguingly, upregulated was also an isocitrate dehydrogenase gene (idh-1). This produces reducing equivalent (NADPH) carrying electrons that may fuel complex I (Smolková et al., 2012; Martínez-Reyes et al., 2020), but it might also add to the stimulation of the antioxidant capacity or to the maintenance of redox homeostasis by regenerating reduced glutathione (Hermes-Lima and Zenteno-Savin, 2002; Penkov et al., 2015; Yang et al., 2019).
If glycolysis is a key process for ATP generation in anoxia (Lutz et al., 1997; Semenza et al., 2001; Hochachka et al., 2001; Huang et al., 2008; Larade et al., 2009) and if, consistently, hxk-2 was upregulated under this condition (Figure 6), based on the expression levels of transcripts encoding for alpha-amylases (see Carbohydrate metabolism in Figure 6), starch and/or glycogen (Jackson and McLaughlin, 2009) may be the prominent carbon sources under anoxic sulfidic conditions.
Ubiquitin-proteasome system and proteases. Proteolysis supplies amino acids or polypeptides to the cells, while impeding the accumulation of damaged or misfolded proteins. The two main mechanisms of cellular proteolysis are the lysosome-mediated intracellular protein degradation (autophagy) and the proteasome-mediated proteindegradation (ubiquitin-proteasome system, UPS). In the latter, ubiquitin-protein ligases covalently attach ubiquitin to proteins, allowing their recognition and further degradation by the proteasome (Lodish et al., 2008; Papaevgeniou and Chondrogianni, 2014).
As shown in Figure 1, transcripts encoding for polyubiquitin (ubq-1), had the highest median gene expression across all transcriptomes. However, all ubiquitination-related genes detected in the differential gene expression analysis between the AS and H conditions, were upregulated in AS worms (Figure 2 and 3, Data S1). For example, aos-1, encoding for a subunit of the ubiquitin-activating enzyme (E1) (Jones et al., 2001), two ubiquitin-protein ligases (E3s without detected cullin domains; Papaevgeniou and Chondrogianni, 2014), and kelch-like genes (e.g., kel-8-like and kel-20). The former are BTB-domain containing proteins known to interact with E3 enzymes, with kel-8 being involved in the degradation of glutamate neuroreceptors (Schaefer and Rongo 2006; Stogios et al., 2005; Kim et al., 2018). Additional ubiquitination-related genes upregulated in AS were csn-2, encoding for a component of the COP9 signalosome complex (Pintard et al., 2003; Brockway et al., 2014), and proteasome genes (pas-2 and pas-3; Fraser et al., 2000; Blumenthal et al., 2002).
Among the proteases that were upregulated in AS worms, aspartyl proteases have been involved in neurodegeneration (Syntichaki et al., 2002), whereas plasminogen and the zinc matrix metalloproteinase ZMP-2 were both reported to mediate degradation of extracellular matrix (ECM) (Vassalli et al., 1991; Altincicek et al., 2010; Fischer, et al., 2014) (Figure 3). C. elegans ZMP-2 was also shown to prevent the accumulation of oxidized lipoproteins (Fischer et al., 2014), and, therefore it may contribute to the enhanced antioxidant response observed in this condition.
Autophagy and amino acid degradation. Besides acting coordinately to withstand stress, autophagy cooperates with apoptotic UPS for the recovery and supply of nutrients when these are scarce (Vabulas et al., 2005; Scott et al., 2004; Huber and Teis, 2016; reviewed in Wang RC et al., 2010 and Russel et al., 2014). Transcripts of two autophagy-related genes, bec-1 (Liang et al., 1999) and the Ragulator complex protein LAMTOR4 (C7orf59-like) (Bar-Peled et al., 2012) were more abundant in AS nematodes (Figure 3). While the former positively regulates autophagy (Liang et al., 1999; Meléndez et al., 2003), the latter interacts with the mTOR Complex I (mTORC1), and tethers small GTPases (Rags and Rheb) to the lysosomal surface (Bar-Peled et al., 2012). When amino acid levels are low, mTORC1 is not translocated to the lysosomal surface (Wang et al., 2009; Bar-Peled et al., 2012), thereby favoring catabolic processes such as autophagy (Thompson et al., 2005). We propose that amino acid scarcity might result from the upregulation of genes involved in the degradation of lysin, glycin, tyrosin, cystein, leucin, isoleucin, valin or tryptophan (Figure 3, Data S1). This would decrease mTORC1 activity and, in turn, stimulates nutrient recycling via autophagy in AS worms.
Conversely, we hypothesize that in H worms, active mTORC1 interacts with the ribosomal protein S6 kinase (S6K), encoded by the rsks-1 gene which is also up in H worms (Ladevaia et al., 2014) (Figure 3). This direct interaction, upon a cascade of phosphorylation events, would stimulate translation, and ultimately cell growth and proliferation (Ma et al., 2009, Howell et al., 2011, and Ladevaia et al., 2014).
All in all, although it is currently unclear whether increased autophagy is beneficial or detrimental, under AS conditions, the upregulation of genes involved in self-digestion might play a protective role and foster recovery from starvation (Thompson et al., 2005), pathogens (Huber and Teis, 2016) or from neuronal and muscular degeneration induced by oxygen deprivation (Murphy and Steenbergen 2008).
Lectins and mucins. Given that symbiont attachment may be mediated by Ca2+-dependent lectins (Nussbaumer et al. 2004, Bulgheresi et al., 2006, 2011) and given that, under anoxia, the symbiont appeared to proliferate more (Paredes et al., 2021), we expected nematode lectins to be upregulated under this condition. Indeed, nine C-type lectin domain (CTLD)-containing proteins were upregulated in AS L. oneistus adults and only two (clec-78 and clec-78-like-2) were upregulated in the presence of oxygen (Figure 4). In addition to CTLD-containing proteins, mucins, a class of glycoproteins with more than 50% of its mass attributable to O-glycans, were also upregulated in AS nematodes. Considering that mucin glycans are used by vertebrate gut commensals for attachment, as well as a source of nutrients (Koropatkin et al., 2012), it is conceivable that their upregulation in anoxia (Figure 4), together with that of CTLD-containing proteins, would foster symbiont attachment.
We hypothesize that overexpression of two classes of putative symbiont-binding molecules, lectins and mucins, under conditions favoring symbiont proliferation (i.e., AS condition, Paredes et al., 2021) may mediate bacterial coat reinforcement.
Apoptosis. Mitochondria play an important role in apoptosis induction (Simon et al., 2000; Martínez-Reyes et al., 2020). Indeed, MPTP opening due to ROS (or the severe ATP decline imposed by the absence of oxygen) may cause cytochrome C release from mitochondria and this, in turn, triggers caspase activation (Martinou et al., 2000; Simon et al., 2000; Gogvadze et al., 2006; Galli et al., 2014). We observed that transcripts encoding for sco-1, a gene needed for the synthesis and assembly of mitochondrial cytochrome C (Williams et al., 2005) were more abundant in AS worms (Figure 4). Further, we observed upregulation of Caspase-3 (ced-3) which belongs to a family of cysteine proteases involved in apoptosis (Mangahas et al., 2005; Kaufmann et al., 2008) and which is activated upon mitochondrial cytochrome C release into the cytosol (Liu et al., 1996; Tafani et al., 2000; Kaufmann et al., 2008; Martínez-Reyes et al., 2020). Additional apoptosis-related genes that appeared to be upregulated in AS worms were: bec-1 (Figure 3), a gene that promotes autophagy and fine-tunes the Ced-3-mediated apoptosis (Liang et al., 1999; Takacs-Vellai et al., 2005); ttr-52, which mediates apoptotic cell recognition prior to engulfment (Wang, X. et al., 2010; Chen et al., 2013); a BAG family molecular chaperone regulator 1 (BAG1-regulator); a cell-death-related nuclease crn−2 (Parrish et al., 2003; Samejima et al., 2005) and phagolysosome forming arl-8 (Sasaki et al., 2013), and a tyrosine kinase Abl-1, (abl-1) that modulates apoptotic engulfment pathways (Hurwitz et al., 2009).
Lipid catabolism. Genes involved in lipid metabolism were similarly expressed between the AS and H conditions (Figure 2, Data S1).In accordance, lipidomes of nematodes incubated in the presence or absence of oxygen were not significantly different (Figure S5, Supplemental material). However, in line with the overall upregulation of degradation pathways, we observed upregulation of genes involved in FA beta-oxidation (kat-1; Berdichevsky et al., 2010), in lipid digestion (the lipase lipl-6; UniProtKB E2S7J2), and lipid degradation (a peripilin-2-like protein; Chughtai et al., 2015). Moreover, a gene that might be involved in oxidative-stress tolerance (a stearic acid desaturase fat-7 regulating the first step of the fatty acid desaturation pathway (Horikawa et al., 2009) was also upregulated in AS worms. Lipid degradation under anoxia might be a strategy to overcome starvation (Krivoruchko and Storey, 2015).
Notably, we also observed an upregulation of two genes involved in phosphatidylcholine (PC) synthesis (pmt-1, pmt-2, Brendza et al., 2007) (Figure 5). Intriguingly, PC was more abundant in the anoxic symbiont (Paredes et al., 2021), although the latter cannot synthetize it. Thus, their upregulation in AS worms suggests worm-symbiont lipid transfer.
GABA- and glutamate-mediated neurotransmission. Upregulated genes related to GABA synthesis were, unc-25, unc-104 and pdxk-1 (pyridoxal phosphate hexokinase) (Thomas et al., 1990; Mclntire et al., 1993; Jin et al., 1999; Gally et al., 2003; Nordquist et al., 2018; Risley et al., 2016) (Figure 5, Data S1). Consistent with an expected increase in glutamate requirement as a direct GABA precursor (Martin et al., 1993), we observed downregulation of two glutamine synthetases and a delta-1-pyrroline-5-carboxylate synthase (gln-3 and alh-13 respectively; van der Vos et al., 2012; Yen et al., 2021; Figure 6), known to convert glutamate to glutamine or to proline, respectively. Furthermore, an mgl-2 like gene encoding for a glutamate receptor, which is activated in the presence of glutamate (Tharmalingam et al., 2012), was up in AS worms. Note that, when oxygen is limited, glutamate may act as a neurotoxic amino acid (Baker et al., 1991; Lutz et al., 2003a). Therefore, increased GABA biosynthesis might, beneficially, prevent its accumulation (Milton et al., 2002; Mathews et al., 2003).
GABA-mediated neurotransmission has been documented for facultative anaerobic animals thriving in anoxic conditions (Lutz et al., 1997; Milton et al., 1998; Lutz et al., 2003a, b). Due to its inhibitory nature, it contributes to avoid membrane depolymerization (Nilsson et al., 1990; Milton et al., 1998). Moreover, given that it relaxes muscles, the increment of GABA may impact the movement of the animal (Mclntire et al., 1993; Schuske et al., 2004). Therefore, upregulation of GABA-mediated neuronal activity might explain why anoxic L. oneistus did not form tight worm clusters after 24h (Supplemental movie 3).
Dopamine-mediated neurotransmission. A gene encoding for the tyrosine hydroxylase Cat-2 (cat-2), which is needed for dopamine biosynthesis (Sawin et al., 2000) and two putative dopamine receptors (protein-D2-like and a G_PROTEIN_RECEP_F1_2 domain-containing protein (dop-5); Sanyal et al., 2004) were upregulated in AS worms. Moreover, a dat-1-like gene mediating dopamine reuptake into the presynaptic terminals was downregulated (Gainetdinov et al., 2002; McDonald et al., 2006) in AS worms (Figure 5).
Calcium-binding and -sensing proteins. Finally, in AS worms several calcium-binding or -sensing proteins (e.g., ncs-2, cex-2, and a calbindin-like (CALB1 homologue); Soontornniyomkij et al., 2012; Hobert et al., 2018; Figure 5), as well as calcium transporters (cca-1, Steger et al., 2005; Transport category, Figure 6) were upregulated. On the one hand, we hypothesize their involvement in the inhibitory neural signaling described above (for example, Ncs-2 mediates the cholinergic and GABAergic expression of C. elegans (Zhou et al., 2017). On the other, they may protect cells against the stress inflicted by anoxia, which involves calcium overload and consequent cellular acidification (Bickler et al., 1992; Dell'Anna et al., 1996; Galli et al., 2014).
Genes upregulated in hypoxic (H) nematodes
Innate immune pathways and effectors. Animals recognize and respond to microbes by means of immunoreceptors including Toll-like receptors, conserved from sponges to humans (Akira et al., 2006). We identified almost all genes belonging to this pathway, including the one encoding for the NF-kB transcription factor. This came as a surprise given that, up to now, the has not been identified in any other nematode NF-kB (Pujol and Ewbank, submitted). As surprising, was the fact that not only two Toll-like receptors (tol-1 and tol-1-like), but also genes encoding for antimicrobial proteins such as a peroxisome assembly factor involved in defense against Gram- (prx-11-like, Wang, D. (2019), a putatively antifungal endochitinase (Dravid et al., 2015) and Bactericidal Permeability Increasing proteins (BPIs) were also more abundant in H worms. BPIs may bind LPS and perforate Gram- membranes and have shown to play a symbiostatic role in other invertebrates (Bruno et al., 2019; Krasity et al., 2015; Chen et al., 2017). However, it is unclear whether activation of the L. oneistus Toll pathway leads to the nuclear NF-kB switching on the expression of antimicrobial genes or whether, as shown in C. elegans, the Toll pathway mediates behavioral avoidance of pathogens (Pradel et al., 2007; Brandt et al., 2015).
Overall, the apparent oxygen stimulation of a central innate immunity pathway and, directly or indirectly, of broad range anti-defense mechanisms could be adaptations to the fact that in oxygenated environments (when crawling in superficial sand layers), L. oneistus is exposed to predation from bigger animals, but also to pathogenic members of the bacterioplankton. Overexpression of broad-range antimicrobials in response to oxygen might therefore help L. oneistus to avoid colonization by potentially deleterious, fouling bacteria (e.g., Vibrios, Roseobacters and Pseudoaltermonas/Alteromonadales) when crawling close to the water column (Dang and Lovell, 2016; M. Mussmann, personal communication).
Development. Although development-related genes were some of the most expressed under all conditions (Figure 1), many were upregulated in H nematodes (Figure 2 and 5). Among the development-related genes upregulated in H nematodes were those related to molting (e.g., nas-36, nas-38, chs-2, ptr-5, ptr-18, apl-1, myrf-1;Suzuki et al., 2004; Zhang et al., 2005; Zugasti et al., 2005; Hornsten et al., 2007; Russel et al., 2011), germ line establishment (e.g., ccm-3, rsks-1;Pan et al., 2007;Pal et al., 2017), oogenesis/spermatogenesis (crt-1,Park et al., 2001), embryonic development and yolk production (smp-1, cpna-1, plt-1, vit-6, crt-1, arrd-17, mlc-5; Clark et al., 1997;Goedert et al., 1996; Gatewood et al., 1997; Fuji et al., 2002; Gally et al., 2009; Zahreddine et al., 2010; Jee et al., 2012; Warner et al., 2013; Fisher et al., 2014; Perez and Lehner, 2019), and/or larval development (nmy-1, ifb-1; Ding et al., 2004;Osório et al., 2019), as well as male tip (Cdt1, plx-1, ver-3, ; Nelson et al., 2011; Dalpé et al., 2004; Dalpe et al., 2013), vulva morphogenesis (hda-1, unc-62), and a hermaphrodite-related gene (hda-1; Dufourcq et al., 2002; Choy et al., 2007) (Figure 5). Morever, transcripts encoding for a number of proteases shown to be involved in C. elegans molting (e.g., nas-38, nas-6-like; Park et al., 2010), development (e.g., teneurin-a-like; Topf and Drabikoswki, 2019), neuronal regrowth or locomotion (tep-1; Kim et al., 2018) and pharingeal pumping (e.g., neprilysin nep-1; Spanier et al., 2005) were also more abundant in H worms. Remarkably, vav-1, which, besides being involved in male tip and vulva morphogenesis (Nelson et al., 2011), may also regulate the concentration of intracellular calcium (Norman et al., 2005), was one of the few development-related genes to be downregulated in H nematodes (see previous section on Ca-binding proteins).
To sum up, and as expected, the host appears to exploit oxygen availability to undertake energetically costly processes, such as development and molting (De Cuyper and Vanfleteren 1982; Uppaluri and Brangwynne 2015).
Carbohydrate metabolism. If in AS nematodes, glycogen or starch appeared prominent carbon sources, H worms seemed to exploit trehalose and cellulose instead. Indeed, genes that degrade trehalose (tre-1, Pellerone et al., 2003) and cellulose (Ppa-cel-2, Schuster et al., 2012) were upregulated in H worms, as well as a putative ADP-dependent glucokinase (C50D2.7) involved in glycolysis (Yuan et al., 2012). The use of this pathway was supported by the overexpression of four genes encoding for sugar transporters (Slc2-A1, C35A11, K08F9.1, F53H8.3; Kitaoka et al., 2013; Bertoli et al., 2015), perhaps switched on by active mTOR (see above) (Figure 6) (Howell et al., 2011).
Additionally, L. oneistus appeared to exploit oxygen to synthesize complex polysaccharides, such as heparan sulfate (hst-1-like; Miyagawa et al. 1988; Bhattacharya et al., 2009) and glycan (Gcnt3-like) (Figure 6), as an ortholog of the N-deactetylase/N-sulfotransferase hst-1, related to heparin biosynthesis was also upregulated (Bhattacharya et al., 2009).
Although glycolysis seems to generate ATP in both AS and H worms, it is not clear why the latter would prefer to respire cellulose or trehalose instead of starch. Given its role as a membrane stabilizer, we speculate that AS worms might prioritize the storage of trehalose over its degradation to preserve membrane integrity (Figure 6) (Crowe et al 1987; Carpenter et al., 1988; Clegg et al., 1997; Chen et al., 2002; Haddad 2006). Of note, based on its genome draft, the symbiont may synthetize and transport trehalose, but it may not use it (Paredes et al., 2021). Therefore, we hypothesize symbiont-to-host transfer of trehalose under hypoxia. Consistently, the symbiont’s trehalose synthesis-related gene (otsB; Paredes et al., 2021), and the host trehalase (tre-1;Figure 6) were both upregulated under hypoxia and metabolomics could detect trehalose in both partners (Table S1). Metabolomics also detected sucrose in both the holobiont and the symbiont fraction (Table S1). Given that, based on transcriptomics and proteomics, the nematode can utilize sucrose but cannot synthesize it (Data S1), whereas the symbiont can (Paredes et al., 2021), as in the case for trehalose, we hypothesize symbiont-to-host sucrose transfer.
Acetylcholine-mediated neurotransmission. Instead of upregulating genes involved in inhibitory (GABA and dopamine-mediated) neurotransmission, hypoxicworms appeared to use excitatory acetylcholine-mediated neurotransmission as indicated by the upregulation of molo-1, acr-20, cup-4, lev-9, and sphingosine kinase sphk-1 that promotes its release(Mongan et al., 2002; Patton et al., 2005; Gendrel et al., 2009; Boulin et al., 2012; Chan et al., 2012) (Figure 5). On the one hand, acetylcholine-mediated neurotransmission might promote ROS detoxification in H worms (Sun et al., 2014). On the other hand, its downregulation in AS worms may beneficially decrease calcium influx (Hochachka and Lutz, 2001).
Feeding, mating, mechanosensory behavior and axon guidance and fasciculation. Transcripts related to the neuronal regulation of energy-demanding activities such as feeding, mating, motion, as well as nervous system development were more abundant in H nematodes (Figure 5, and Data S1). More precisely, upregulated genes were involved in pharyngeal pumping (nep-1, lat-2;Spanier et al., 2005; Guest et al., 2007), male mating behavior and touch (pdfr-1, tbb-4, ebax-1, Hurd et al., 2010; Wang, Z. et al., 2013), axon guidance and fasciculation (spon-1, igcm-1, ebax-1, tep-1; Kim et al., 2018; Woo et al., 2008; Schwarz et al., 2009;Wang, Z et al., 2013), mechanosensory behavior (e.g., mec-12, delm-2; Gu et al., 1996; Han et al., 2013). Additionally, we also observed the upregulation of a gene encoding for a glutamate receptor (glr-7) possibly involved in feeding facilitation (Li et al., 2012).
Amino acid biosynthesis. Transcripts of genes involved in the synthesis of glutamine and proline (gln-3 and alh-13, respectively),aspartate (L-asparaginases; Tsuji et al., 1999) and S-adenosyl-L-methionine (SAM) (sams-4; Chen et al., 2020) were all upregulated in H worms (Figure 6), as well as one encoding for the ornithine decarboxylase odc-1 which is involved in biosynthesis of the polyamine putrescin, and is essential for cell proliferation and tissue growth (Russell et al., 1968; Heby, 1981). Moreover, polyamines, with their high charge-to-mass ratio may protect against superoxide radicals, which, as mentioned, harm cell membranes and organelles, oxidize proteins, and damage DNA (Gilad et al., 1991; Longo et al., 1993).
Lipid biosynthesis. Genes upregulated in H worms mediate the biosynthesis of long chain fatty acids (acs-3, acs-14, elo-3 but not acs-5;Yuan et al., 2012; Ward et al., 2014; Wang et al., 2021), sphingolipids (a sphingosine kinase-1 (sphk-1) and egl-8, which controls egg laying and pharyngeal pumping in C. elegans (Bastiani et al., 2003). Notably, sphingolipids may be anti-apoptotic (Taha et al., 2006) or result in acetylcholine release (Chan et al., 2012).
On the other hand, ceramides, which have antiproliferative properties and who may mediate resistance to severe oxygen deprivation (Deng et al., 2008; Menuz, et al. 2009), appeared to be mainly synthesized in AS worms, as indicated by the upregulation of genes involved in ceramide biosynthesis (asm-3, ttm-5; Watts et al., 2017) (Figure 6).
Transport. As anticipated in the introduction, anoxia-tolerant animals switch off ATP-demanding processes such as ion pumping (Lutz et al., 1996; Galli et al., 2014). Indeed, transcripts encoding for proteins involved in cation channel activity (gtl-2, voltage gated H channel 1; Teramoto et al., 2010), sodium transport (delm-2-like; Han et al., 2013), chloride transport (anoh-1, best-13, best-14; Tsunenari et al., 2013; Wang, Y. et al., 2013; Goh et al., 2018), ABC transport (wht-2, pgp-2, slcr-46.3, F23F12.3, hmit-1.3; Currie et al., 2007; Schroeder et al., 2007; Kage-Nakadai et al., 2011) and organic transport (F47E1.2, oct-2; Pao et al., 1998) were all more abundant in H than AS worms (Figure 6).
Sulfur metabolism. The mpst-7 gene which is involved in organismal response to selenium and it is switched on in hypoxic C. elegans (Romanelli-Credrez et al., 2020) was upregulated in H nematodes (Figure 6). Given that the latter is thought to catalyze the conversion of sulfite and glutathione persulfide (GSSH) to thiosulfate and glutathione (GSH) (Filipovic et al., 2018), hypoxia-experiencing L. oneistus might express this enzyme to recharge the cells with GSH and hence, help to cope with oxidative stress (Hayes and McLellan, 1999; Mytilineou et al., 2002; Diaz-Vivancos et al., 2015). Also more abundant in H worms were transcripts encoding for the sulfatases 2 (sul-2) (Morimoto-Tomita et al., 2002) and a PAPS-producing pps-1 (3′-phospho-adenosine-5′-phosphosulfate (PAPS) considered the universal sulfur donor; Bhattacharya et al., 2009), as well as for the chaperones pdi-6 and protein-disulfide-isomerase-A5-like which require oxygen to mediate correct disulfide bond formation in protein folding (Teodoro and O’Farrell, 2003; Rose et al., 2017; Livshits et al., 2017) (Figure 6).
Conversely, a putative sulfide-producing enzyme (mpst-1) who protects C. elegans from mitochondrial damage (Qabazard et al., 2014; Ng et al., 2019; Kimura, 2020) was upregulated in AS nematodes. Notably, under AS, L. oneistus might detoxify sulfide by producing glutathione and taurine (Rose et al., 2017), as a persulfide dioxygenase (ethe-1) and a cysteine dioxygenase (cdo-1) which catalyzes taurine synthesis via cysteine degradation were upregulated. Sulfide detoxification via taurine accumulation is a common strategy in chemosynthetic animals (reviewed in Cavanaugh et al., 2006).
All in all, L. oneistus appeared to limit excess accumulation of free sulfide in anoxia and to free sulfate when oxygen was available.
{kind=link}