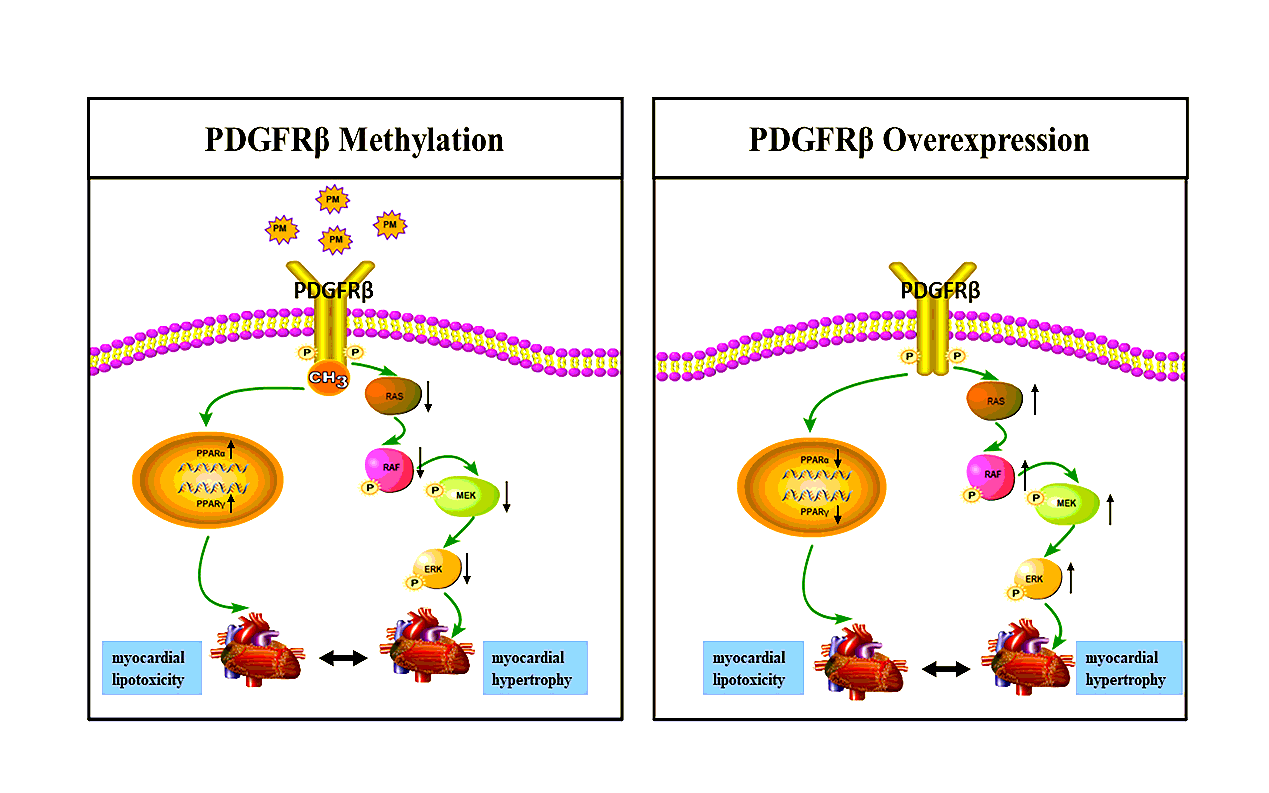
1. PM induced epigenetic alteration and DNA methylation profiling identified the key gene-PDGFRβ in heart tissue of C57BL/6J mice.
In order to explore the epigenetic changes that occur in mouse hearts after a real-ambient PM exposure via an individual ventilated cage (IVC)-based system, we performed reduced representation bisulfite sequencing (RRBS). As can be seen from the distribution map of the number of methylation levels in functional regions, the genes with hypermethylation or hypomethylation were mainly concentrated in intron regions, while the difference between high and low methylation was the largest in repeat region (Fig. 1a). According to the length distribution map of differential methylation region (DMR), the length of DMR is between 0 and 400bp (Fig. 1b). Although the difference in discrete values between the exposed group and the control group was not significant, the distribution of DMRS methylation level in the exposed group was relatively concentrated (Fig. 1c). The heat map analysis of the methylation level in the functional region between the samples showed that there was significant difference methylation between the exposed group and the control group. Among them, there were 3808 (57.66%) hypermethylated genes and 2796 (42.34%) hypomethylated genes (Fig. 1d). GO and KEGG analysis revealed the GO terms and important KEGG pathways which PM may trigger. According to GO analysis results, in the three aspects of biological process, cell component and molecular function, differentially methylated genes mainly concentrated in the growth and development process of biological process, the development process of anatomical structure, single-multicellular biological process and other categories. In KEGG result analysis, differentially methylated genes mainly regulated the PI3K-Akt signaling pathway, RAS signaling pathway, MAPK signaling pathway and Rap1 signaling pathway. (Fig. 1e and Fig. 1f). Interestingly, the differential methylated gene PDGFRβ in this study was enriched in the MAPK pathway and was associated with multiple pathways. We found that it is closely associated with the downstream MEK/ERK pathway. Overall, we found that PM induced epigenetic changes and RRBS revealed a hypermethylated gene PDGFRβ. Given the established roles of PDGFRβ signaling, we investigated the effects of PDGFRβ and its downstream related pathways in our subsequent experiments.
2. PM induces PDGFRβ hypermethylation mediated MEK/ERK pathway to trigger myocardial hypertrophy in heart tissue of C57BL/6J mice.
Exposing mice to 6 weeks of real-ambient PM via the IVC-based system resulted in a significantly thickened right ventricular wall in mice (Fig. 2a, b, c). To verify the methylation status of PDGFRβ, in the following RT-qPCR analysis, compared with the control group, the mRNA expression level of PDGFRβ in the PM exposure group showed a significantly decreased trend, which was also consistent with the results of the RRBS analysis (Fig. 2d). Western blot analysis also showed that the protein levels of RAS, RAF, MEK and ERK, which are downstream of PDGFRβ via the MEK/ERK pathway, showed a downward turn in the PM exposure group compared to control (Fig. 2e, f, g, l). Hence, our results indicate that PM directly leads to cardiac hypertrophy and hypermethylation of PDGFRβ in mice, which leads to down-regulation of the expression levels of related genes in its downstream MEK/ERK pathway.
MEK/ERK pathway is closely related to cardiac hypertrophy. In order to further understand the mechanism of MEK/ERK pathway induced cardiac hypertrophy after PDGFRβ hypermethylation, primary neonatal rat cardiomyocytes were exposed to different concentrations of PM. CCK8 assay showed a stress-induced increase in cell viability at 25 μg/mL, and then showed a dose-dependent decrease with the increase of PM concentration (Fig. 3a). Finally, we selected three different PM concentrations for further study. The results of RT-qPCR showed that the mRNA expression levels of PDGFRβ and downstream RAS, RAF, MEK and ERK all decreased with increased PM concentrations (Fig. 3b, c, d, e, f). Importantly, these results were consistent with western blot analysis of protein expression (Fig. 3g, h, j, k, l). Based upon these data, we chose 50μg/ mL PM as our experimental concentration for future experiments. However, we also know that PM exposure time has a significant effect on experimental results. To assess this, we established PM exposure models at different time points in primary neonatal rat cardiomyocytes to detect the expression of myocardial hypertrophy markers. RT-qPCR results showed that mRNA levels of cardiac hypertrophy related markers in cardiomyocytes showed an upward trend during the first 0-6 hours of PM exposure. This was followed by a decrease in mRNA expression during the 6-24 hours timepoints (Fig. 3m, n, o). Based upon these results, we conducted the study after 6 hours of PM exposure. We found that when PM concentration was 50μg/ mL and exposure time was 6 hours, the mRNA expression levels of the three DNA methyltransferases followed an upward trend (Fig. 3p, q, r). Thence, we conclude that exposing cardiomyocytes to PM at 50 μg/mL for 6 hours causes changes in DNA methylation levels in cardiomyocytes and may affect cardiac hypertrophy.
3. Effects of up-regulation of PDGFRβ on MEK/ERK pathway and cardiac hypertrophy in cardiomyocytes.
To investigate the effect of PDGFRβ on MEK/ERK pathway and cardiac hypertrophy. We used a recombinant enhanced green fluorescent protein (EGFP) plasmid with high expression of PDGFRβ (hPDGFRβ) and an empty plasmid (PDGFRβ-con) for transient gene transfection in cardiomyocytes after PM exposure. Fluorescence images demonstrate that the transfection was successful (Fig. 4a). Then we performed RT-qPCR and WB to verify that overexpression was achieved. The mRNA level of hPDGFRβ was 28 times higher than that of blank control and PDGFRβ-con (Fig. 4b). The expression level of the protein was also consistent with that of RT-qPCR (Fig. 4c). We found that RT-qPCR results showed that mRNA expression levels of RAS, RAF, MEK and ERK were significantly decreased in the PM exposure group compared with the control group, which was consistent with the results of real PM exposure in our previous animal experiments. The mRNA expression levels of RAS, RAF, MEK and ERK in the hPDGFRβ group showed an upward trend (Fig. 4d, e, f, g), and the protein expression levels of MEK and ERK in the WB experiment were also consistent with the results of RT-qPCR (Fig. 4h, I, j). We further verified the effect of hPDGFRβ on cardiac hypertrophy markers by RT-qPCR. We found that the mRNA expression levels of atrial natriuretic factors (ANF), brain natriuretic factor (BNF) and beta-myosin heavy chain (β-MHC) were significantly increased in the PM exposure group, which was consistent with the results of our previous experiment. The mRNA expression levels of ANF, BNF and β-MHC in hPDGFRβ group showed a decreasing trend (Fig. 4k, l, m). These results suggest that high expression of PDGFRβ facilitates increased activation of the MEK/ERK pathway, thereby preventing the increase in cardiac hypertrophy markers, suggesting a net effect ofreducing cardiac hypertrophy.
4. Effect of inhibiting methylation on MEK/ERK pathway and cardiac hypertrophy in cardiomyocytes.
In order to further explore the effects of PDGFRβ methylation on MEK, ERK pathways and cardiac hypertrophy, we established a methylation inhibition model in primary neonatal rat cardiomyocytes. Firstly, CCK8 experiments were conducted to understand the effects of different concentrations of methylation inhibitors (5-Azacytidine: 5AZA) on cell viability (Fig. 5a). According to RT-qPCR results, when the concentration of 5AZA was 10μM, the mRNA expression level of DNA methyltransferase 1(DNMT1) and DNA methyltransferase 3A (DNMT3A) could be decreased (Fig 5b, c), so we chose 10μM as our experimental concentration. We found that the mRNA expression levels of MEK, ERK, RAS and RAF in the methylation inhibitor group showed an upward trend compared with the PM exposure group (Fig. 5d, e, f, g). In the western blot results, the protein expression levels of MEK and ERK were also consistent with the RT-qPCR results (Fig. 5h, I, j). In order to further study the effect of methylation inhibitors on cardiac hypertrophy, we used RT-qPCR to observe the expression levels of markers related to cardiac hypertrophy. We found that compared with PM exposure group, the mRNA expression levels of ANF, BNF and β-MHC in the methylation inhibitor group showed a decreasing trend (Fig. 5k, l, m). These results indicated that the methylation inhibitor group activated the MEK/ERK pathway, reduced the effect of cardiac hypertrophy, and had a protective effect on cardiomyocytes.
5. Effects of up-regulation of PDGFRβ on MEK/ERK pathway and cardiac hypertrophy in heart tissue of C57BL/6J mice.
Since the activation of cardiac-specific transgenic PDGFRβ has a significant effect, we further investigated the regulatory mechanism of high expression of PDGFRβ on cardiotoxicity induced by PM exposure in mice and the potential of PDGFRβ in improving cardiac hypertrophy outcomes. Our adenovirus helper-free system is composed of three plasmids, namely a viral vector, PAAV-RC vector and pHelper vector. We generated AAV9: Ubi by using myocardial cell-specific ubiquitination protein promoter and triple FIAG marker.3Flag-PDGFR β (hPDGFRβ), as a control, we generated AAV9: Ubi-EGFP (hPDGFRβ-Con), and replaced 3Flag-PDGFR β with EGFP (Fig. 6a). AAV9 was first injected into mice by tail vein, and PDGFRβ expression was highly specific in the heart after 20 days. The high expression of PDGFRβ was clearly observed under the light microscope by immunohistochemical experiments, and the green fluorescence of AAV9 in the heart tissue was directly observed by frozen sections, which was consistent with the immunohistochemical results (Fig. 6c). In addition, we also examined the expression level of PDGFRβ protein by western blot assay, and we found that the protein expression level of PDGFRβ group was higher than that of hPDGFRβ-con group (Fig. 6d), so 20 days later, the PDGFRβ high expression model was established. Then we performed tracheal drip, once a week, where the PM of tracheal drip came from IVC-based real-ambient PM exposure system, and the total concentration of tracheal drip was consistent with the total concentration in real-ambient PM exposure system (Fig. 6b). Then we conducted echocardiographic examination of the mice, and it was found that interventricular septum (IVS), interventricular septal thickness at systolic (IVSs), interventricular septal thickness at diastole (IVSd), left ventricular posterior wall (LVPW), left ventricular posterior wall of systolic (LVPWs), left ventricular posterior wall of diastolic (LVPWd) in the PM exposed group showed an increasing trend compared with the control group, indicating that myocardial hypertrophy appeared in the exposed group. Compared with PM exposure group, hPDGFRβ group showed a decreasing trend in all indicators (Fig. 6e, f, g, h, i, j), indicating that hPDGFRβ can reduce cardiac hypertrophy and thus protect the heart. To follow up these data, we further studied this mechanism. We used western blot experiments to study the related genes in the downstream pathway of PDGFRβ. We observed that RAS, RAF, MEK, pMEK, ERK and pERK in the downstream pathway all showed the negative same trend. The protein expression level showed an upward trend (Fig. 6k, l, m, o, p, q), which was consistent with the results of our previous cell experiments. In conclusion, hPDGFRβ activates downstream MEK and ERK pathways, ameliorates the effects of cardiac hypertrophy and protects the mouse heart.
6. PM - induced myocardial hypertrophy and lipid metabolism disorders.
These data have demonstrated that PM exposure can lead to cardiac hypertrophy in mice. However, whilst is well known that cardiac hypertrophy is closely related to lipid metabolism, we next asked how is PDGFRβ related to lipid metabolism? To further explore the relationship between the two, we first used HE staining to observe the relationship between the PM exposure group and the control group in AC16 cells (Fig. 7a). We found that compared with the control group, the cardiomyocytes in the exposed group showed a decreased nucleo-plasmic ratio (Fig. 7b) and an increased area ratio (Fig. 7c), combined with our previous experimental results in primary neonatal rat cardiomyocytes, indicating that PM exposure did cause myocardial hypertrophy in cardiomyocytes. Next, oil red O staining was used to stain AC16, and we observed that the PM exposure group had significant lipid accumulation (Fig. 7d). Subsequently, oil red O staining was used to observe the heart tissues of the tracheal drip group and the control group, and it was found that the tracheal drip group also had a small amount of lipid accumulation (Fig. 7e).
In summary, PM exposure can lead to lipid metabolism disorder in the mouse heart. Since PPARα and PPARγ are closely related to lipid metabolism, we first used western blot analysis to investigate changes in PPARα and PPARγ protein expression in the heart tissue of real-ambient PM exposure system. We found that the protein expression levels of PPARα and PPARγ in the PM exposure group were increased compared with the control group (Fig. 7f, g, h). Further, PM exposure is closely related to cardiac lipid metabolism. And how does this relate to PDGFRβ? As a result, we established a model of PDGFRβ overexpression in AC16 cells. Using western blot experiments, we found that PPARα protein expression was increased in the PM-exposed group compared with the control group, consistent with previous results, but PPARα protein expression was lower in the hPDGFRβ group than in the exposed group (Fig. 7i, j). Similarly, we also obtained consistent results in the high expression model of PDGFRβ after tracheal drip, with PPARα protein expression increased in the PM exposure group compared with the control group, but decreased in the hPDGFRβ group compared with the exposure group (Fig. 7k, l). In conclusion, hPDGFRβ protects the mouse heart by alleviating PM-induced lipid metabolism disturbances in cardiomyocytes and the mouse heart.
In order to further study the PM induced changes in lipid metabolism, we performed lipidomics analysis using LC-MS high-throughput analysis technology in the tracheal drip mode. Firstly, in the lipid subclass analysis, a total of 14 lipid molecules were detected in the positive ion mode, which were phosphatidylcholine(PC), phosphatidylethanolamine (PE), phosphatidylserine (PS), phosphatidylglycerol (PG), phosphatidic acid (PA), Cardiolipin (CL), sphingomyelin (SM), ceramide (Cer), glucosylceramides (GlcCer), diacylglycerols (DAG), triacylglycerols (TAG), a to carotenoids (ACar), bone morphogenetic protein (BMP) and human bone morphogenetic protein (HBMP). Among them, PC has the most subcategories, up to 348, and TAG has 219 subcategories (Fig. 8a). A total of 16 lipid molecules were detected in negative ion mode, which were PC, PE, PS, phosphatidylinositol (PI), PG, PA, CL, SM, Cer, GlcCer, monogalactosyldiacylglycerol (MGDG), sulfoquinovosyldiacylglycerol (SQDG), fatty acid esters of hydroxy fatty acids (FAHFA), HBMP and ganglioside (GM3). Among them, PC has the most subtypes, with 101 species, followed by PE with 90 species (Fig. 8b). We found that the PC subclass was highest in both positive and negative ion modes. Further analysis of the different lipid compounds revealed that a total of 1226 differential lipid compounds were detected in positive ion mode, among which 34 which were significantly different, 30 which were significantly up-regulated and 4 which were significantly down-regulated. In the negative ion mode, a total of 582 different lipid compounds were detected, among which 38 were significantly different, 29 were significantly up-regulated and 9 were significantly down-regulated. Then, we observed the overall distribution of different lipid compounds from the volcano diagram. It could be seen intuitively that no matter in the positive ion mode or the negative ion mode, more lipid compounds were up-regulated than down-regulated, and the expression multiple of lipid compounds in different groups changed significantly (Fig. 8c, d). So, we then looked at the fold change (FC) analysis of differential lipid compounds and found that in the positive ion mode, Creatine had the largest differential multiple, followed by canrenone. In the negative ion mode, carnosine had the largest differential multiple, followed by L-Histidine (Fig. 8e, f). In order to compare the differences of metabolic expression patterns between two groups and within the same comparison group, hierarchical cluster analysis was performed on the obtained metabolites. We found significant differences between the control group and the exposed group (Fig. 8g, h). In order to check the consistency of lipid compounds and lipid compounds, Pearson correlation coefficient between all lipid compounds was calculated to analyze the correlation between each lipid compound. We found that as the linear relationship between the two lipid compounds increased, the positive correlation tended to 1, and the negative correlation tended to -1(Fig. 8i, j). In general, these data suggest that PM exposure can lead to changes in cardiac lipid metabolism in mice.
{kind=link}