In this study, we investigated the effects of environmental modification on the development, survivorship and fecundity of malaria vector mosquitoes. We hypothesized that irrigated sugarcane plantation area enhances development, survivorship and fecundity compared to non-irrigated field crop area due to better microclimate and nutrients following environmental modification. However, the study showed no significant difference in development and survivorship of An. gambiae s.l. immatures between the two areas.
Variation in vegetation cover may affect the radiation flux and energy balance of the land surface and thus may modify the microclimate [25]. By the time experiment was conducted, sugarcane plantation was at its maturity stage, which is dense and leafy that partly could limit direct sunlight to reach to the washbasins, whereas in the surrounding crops field area were relatively less dense. The mean hourly water temperature in non-irrigated area increased by 1.1 °C compared to irrigated area. This could partly explain the observed 5.1% more pupation rate in non-irrigated area compared to irrigated area. Studies reported elsewhere indicated that an increased temperature due to land use and land cover increased larval survival rate [10, 26–30]. Tuno et al. [29] reported that the survivorship of An. gambiae larvae was reduced from 56% in habitats fully exposed to sunlight to 1.5% in habitats with forest canopy in western Kenya. Wang et al. [10] also reported pupation rate of An. minimus, malaria vector in china to be 52.5%, 12.5% and 3.8% in the deforested, banana plantation and forested areas, respectively, which is far lower than our findings, 79.4% and 84.5% at irrigated and non-irrigated areas, respectively.
Nutrient availability may affect the survival, pupation rate and development time. The potential food source of anopheline larvae may include but not limited to bacteria, fungi, debris and organic matter. The abundance and structure of microbes such as algae and photosynthetic cyanobacteria in aquatic habitats may have changed in response to land use and land cover [31, 32]. Organic matters and debris in the soil at different settings may not be the same, which could possibly vary with change in surrounding land use and land cover. Kebede et al. [33] reported that maize pollen provides nutrition for larval anopheline mosquitoes showing that the incidence of malaria was about 10 times higher in high maize-cultivation areas. In our case, the debris of sugarcane plantation and other field crops might not be the same but the result showed both areas are supporting mosquito development, which needs further investigation of a soil’s biological and chemical composition in relation to mosquito immatures development.
The higher pupation rate and longer survivorship of An. gambiae s.l. immatures generally could increase vectorial capacity to enhance malaria transmission. Based on these findings alone, we cannot conclude that the irrigated area is encountering less malaria risk. Recently, in the study conducted from the same area, significantly more diverse breeding sites and larvae abundance in irrigated sugarcane plantation area than its surrounding has been reported [23]. Thus, more diversified breeding sites with 79.4% pupation rate could certainly outweigh the malaria burden over surrounding environs with less habitat diversity and relatively the same pupation rate.
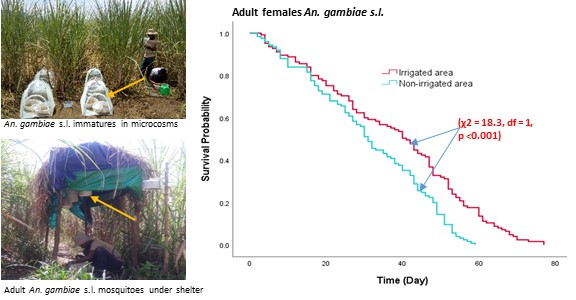
Adult An. gambiae s.l placed in the irrigated area survived longer than non-irrigated area. Adult female mosquitoes survived longer than male in both settings. Our findings of mosquito longevity was in line with previous studies elsewhere. For instance, Okech et al. [34], reported mean survival of 33 days for An. gambiae s.l in western Kenya, which is 6 days shorter than our finding. Gary and Forster [35], found that An. gambiae s.l. mosquitoes had a median survival time of 29 days under insectary conditions, but in our study, the median survival time for female An. gambiae s.l. was 41 and 31 days at irrigated area and filed crop area, respectively. The longer survival of mosquito at the irrigated area indicates that An. gambiae s.l. is well adapted to the environmental conditions. Enhanced survival of malaria vector is among the determinants of increased mosquito vectorial capacity [36]. A long-lived adult female mosquito increases opportunities to encounter an infected human host, the extrinsic incubation period of malaria parasites and reach the salivary glands after an infective blood meal, and transmission of parasites in later blood meals to uninfected hosts [1, 3, 37]. Thus, it has an implication on malaria transmission at the locality.
The experiment set-up at both study settings made the same and human blood and sugar were provided in a similar way. Thus, the only difference was the environment where the experiments were situated, being irrigated area and non-irrigated area. There was no significant difference in mean, maximum and minimum hourly ambient temperature; and relative humidity between two environments. Previous studies indicated that An. arabiensis, a primary vector in Ethiopia, generally prefers areas with low humidity and high temperature [38]. A similar study also demonstrated that reduced humidity and increased temperatures following deforestation creates a more suitable environment for adult An. arabiensis to survive longer [26]. Therefore, in our study setting the determinants involving in supporting better survival of adult An. gambiae s.l. at irrigated warrants further investigation.
The average daily fecundity of An. gambiae s.l. mosquitoes in irrigated area was 96.2% higher compared to non-irrigated area. Increased survival together with enhanced fecundity of malaria vector in irrigated area suggests that the longevity and biotic potential of An. gambiae s.l. in the area is very high, favoring increased population density and thus the species could contribute much to malaria transmission. Better survival and fecundity in the irrigated area in our study is in agreement with the study conducted in Ethiopia at the laboratory level demonstrated that gravid An. arabiensis females attracted to sugarcane pollen volatiles [39].
This study had several limitations. The experiment was done in one time of the maturity stage of irrigated. The microclimate conditions in irrigated area during seedling/ germinating stage, tillering stage, grand growth stage & maturity stage [40] could not be the same which in turn influence the mosquito survivorship. The information on chemical and nutrient’s composition of a soil used as a substrate was not included in the study. Moreover, the experiments were conducted under controlled condition of all potential biological factors that may influence mosquito survival like predators and competitors, which might possibly leads to overestimated survival time than actual.
{kind=link}