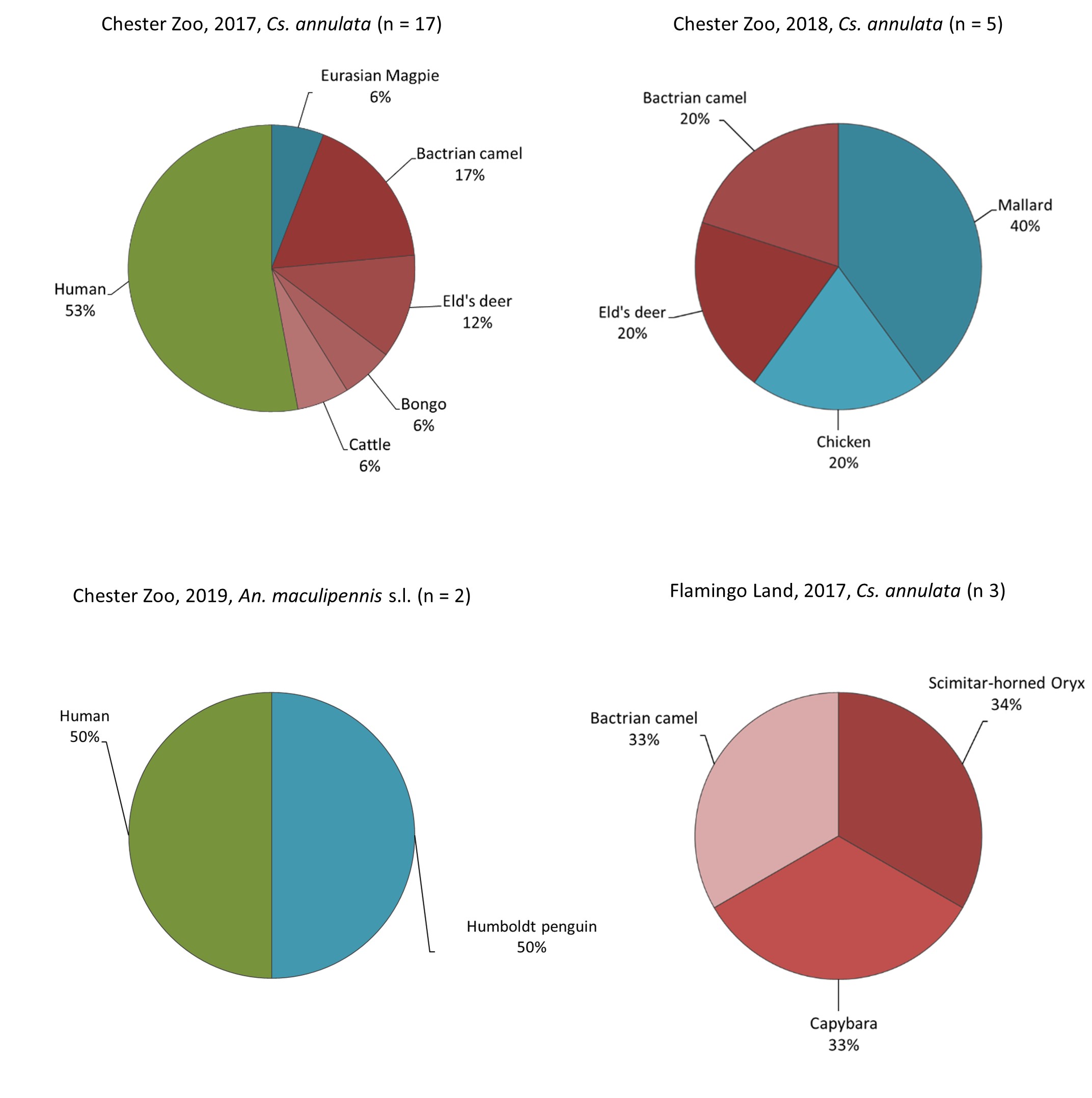
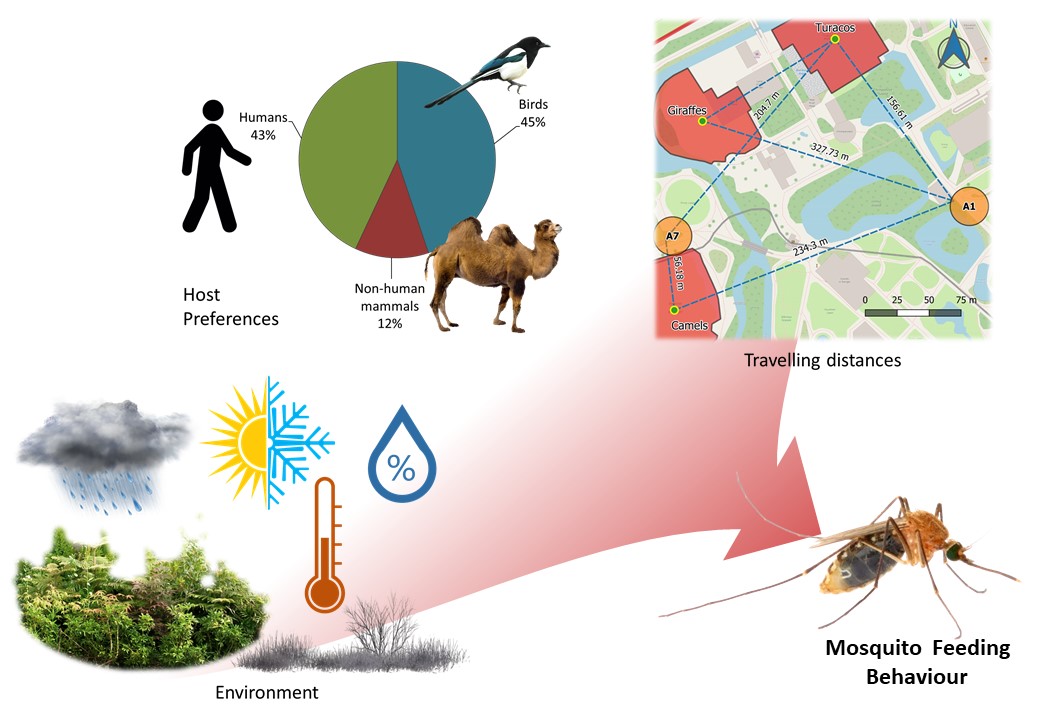
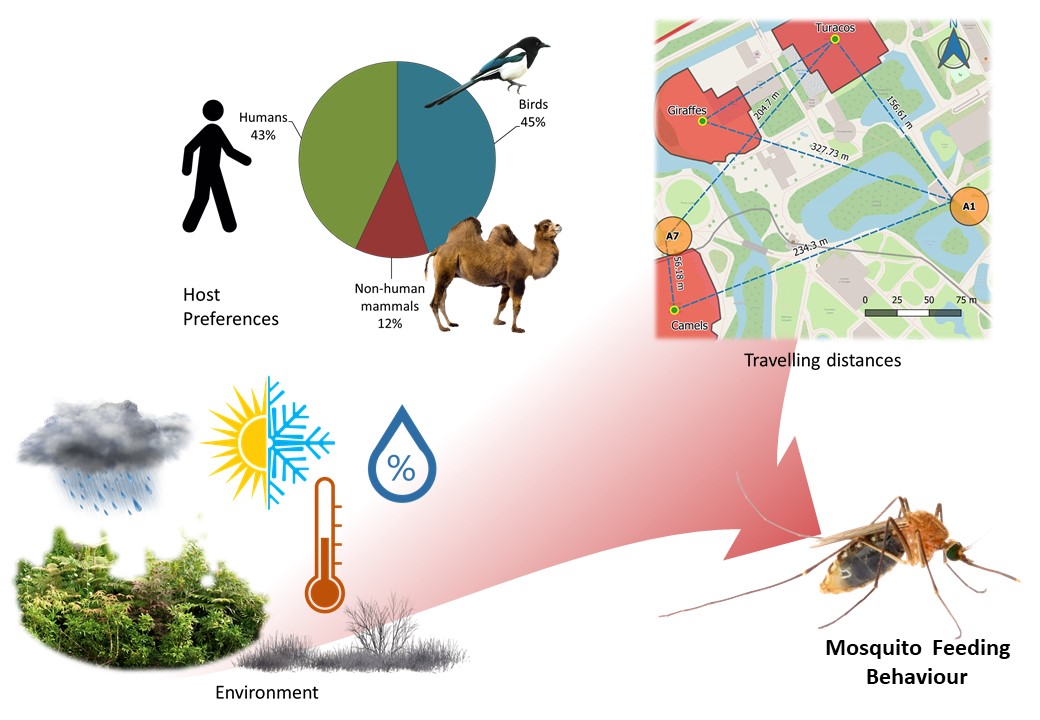
We confirmed the intrinsic preference of Culex pipiens for birds, mainly free wild birds and some birds from the zoos’ collections, and of Culiseta annulata for mammals. Cx. pipiens is primarily an ornithophilic species [35, 40], however we observed high proportions of human hosts in all our samplings, except for 2019; similarly Heym et al. [3] and Börstler et al. [41] found mosquitoes feeding on humans but the proportion we observed was higher overall. We discount the possibility of major sample contamination as negative controls did not produce a positive result and no sequenced samples matched the positive control. These mosquitoes fed on humans from April to August, when the zoos have more visitors and temporary staff, and no human blood-meals were found at other times, although the occurrence of birds and other mammalian blood-meals continued to be observed. Additionally, the preference for humans was significantly higher in Cx. pipiens than in Cs. annulata, which is unexpected as Cs. annulata has been reported as a biting nuisance for people in the UK [42]. PCR identification showed that all Cx. pipiens that fed on humans belong to the pipiens biotype; as this biotype is typically described as ornithophilic, abundance of visitors and staff seem to be a relevant influence in mosquito feeding preferences in zoos.
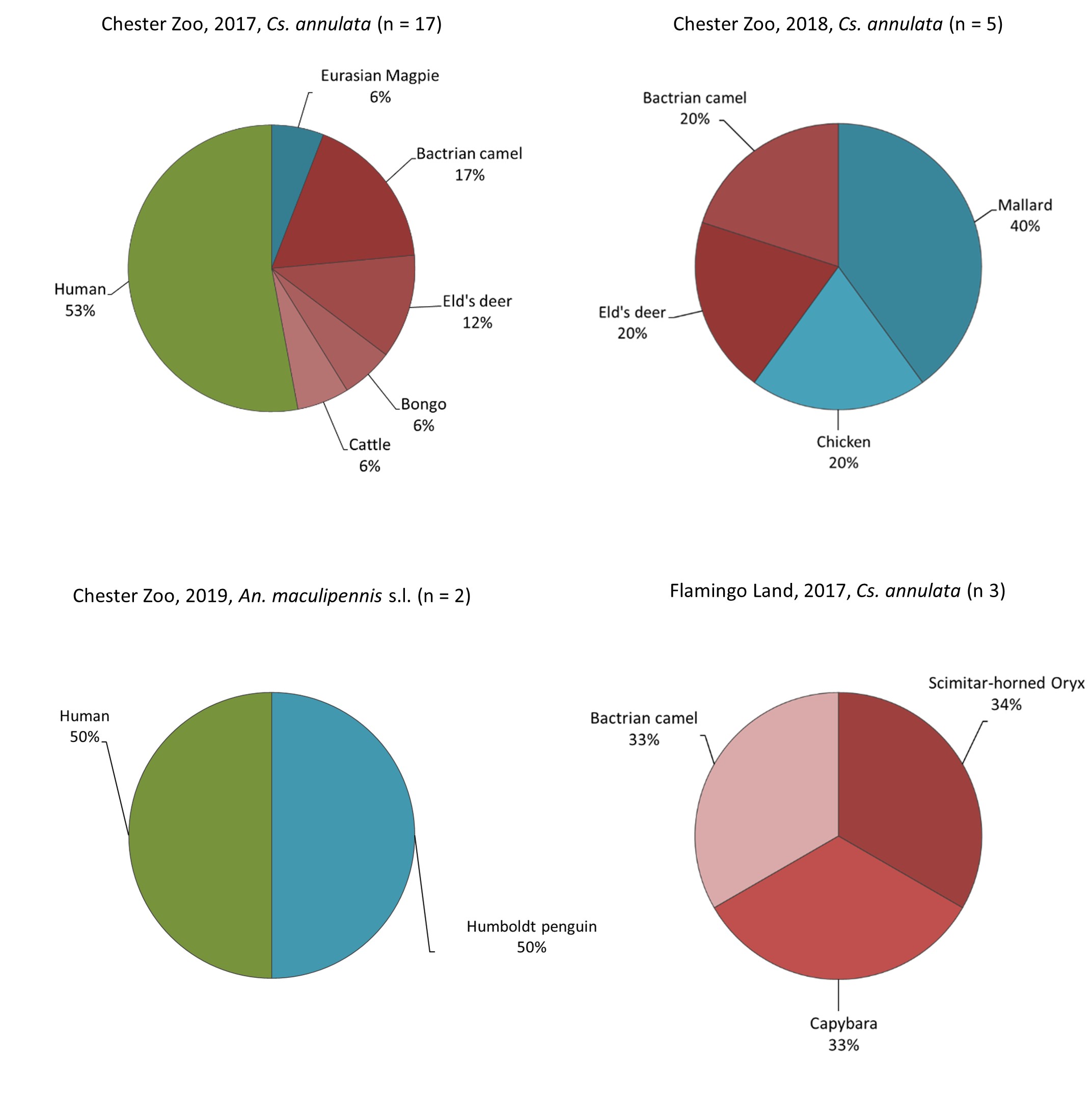
Anopheles maculipennis s.l. prefers to feed on mammals than on birds [32, 43]. However, two mosquitoes from this group fed on Humboldt penguins (Spheniscus humboldti), which to our knowledge is the first report of this host choice. Due to the low sample size, we cannot conclude if there is a host preference or simply a tendency of capturing blood-fed mosquitoes in proximity to the animals they feed on as these mosquitoes were captured close to the penguin exhibit (< 22 m). Therefore, targeted sampling of this group is needed and should include the molecular identification of the species as they have different host preferences [43].
A correlation between mosquito abundance and proportion of blood-feds has been noticed before [21]. During the first year in Chester Zoo, we found high mosquito abundance and high proportion of blood-feds in areas A1 and A3 and again for A1 in 2018. In Flamingo Land, we observed a similar situation in area A2. Nevertheless, other areas with lower general catches showed high proportions of blood-fed mosquitoes, like A7 and A10 in Chester Zoo in 2017 and areas A10 and A11 in 2018. Therefore, the mosquito abundance is not the only explanatory factor. For instance, area A3 in Chester Zoo captured a high number of blood-fed mosquitoes in 2017 possibly due to its proximity to a picnic area and a children’s playground. Fewer mosquitoes were caught in this area in 2018, when the children’s playground was closed for renewal over several weeks, and the off-show aviaries next to the sampling area were expanded, reducing a considerable portion of the vegetation. In Chester Zoo, area A7, which caught more blood-feds than expected in 2017, is also near a picnic garden and an area with high transit of visitors. In 2017 and 2018, area A10 also captured a high proportion of blood-feds which matched mainly wild birds. This area is inside a wetland aviary where several species are kept, and the abundance of wild passerines could be high as they are attracted to the waterfowl’s food despite the netting of the exhibit. In Flamingo Land, area A2, also with a higher proportion of blood-feds, is in the boundary of the South American exhibit which contains large mammals such as capybara (Hydrochoerus hydrochaeris), on which we found two mosquitoes feeding. It appears from this result that the constant presence of suitable hosts is an attractant factor for mosquitoes.
The mosquito host feeding preferences varied each year. There is evidence that host selection by Culex spp. is influenced primarily by the availability of preferred hosts [44]; therefore, these variations possibly depend on the mosquito abundance and host availability, both of which increase during the summer. We observed a significant host shift in the case of Cx. pipiens in Chester Zoo in 2017, preferring humans in June and birds in July. Tuten et al. [2] also observed a host shift for Cx. pipiens, although in this case preferring birds in the summer and birds and mammals in autumn. These changes could be related to host availability influenced for instance, by the migration and breeding seasons of birds (nestlings are more prone to mosquito bites [3]). Furthermore, when the preferred hosts of Culex spp. are scarce, this mosquito can shift to other hosts, including humans [12].
Some species like Cx. pipiens breed and rest close to their host’s habitat [20], so it is reasonable that the chances of capturing blood-fed mosquitoes are higher closer to the location of potential hosts as we observed for zoo animals. Other authors reported maximum travelling distances between 170 to 770 m [2–4, 22] and the maximum that we observed was within this range (367 m). However, we found mosquitoes feeding on domestic animals (cattle, pig, chicken, and dog) that we assume came from nearby farms. Thus, dispersal was probably further than our estimate.
It is possible that in some areas the landscape forms flight paths that aids mosquito’s movement in a certain direction [22] and the dominant wind direction could be a relevant factor influencing dispersal and flight direction [45]. For example, Area A1 in Chester Zoo captured blood-fed mosquitoes that feed on zoo animals located from the southwest to the northwest from this area, and the wind on the day before collection came from a similar direction in the case of four out of six mosquitoes. The use of portable weather stations could improve the study of wind and landscape influence in mosquito dispersal. It is important to consider that using the exhibit’s centroid, or any other measurement to the exhibit, assumes that the animals distribute randomly within their exhibits, when in reality, animals spend more time in certain areas, like around the feeders during the day or in the enclosure at night-time. The estimation of animal occupancy degrees inside the exhibits should be considered for a more precise assessment of mosquito travelling distances.
Temperature affects mosquito feeding activity, reproduction, longevity, and development [46, 47], which could explain the positive relationship with the capture of blood-fed mosquitoes showed in the models. High relative humidity may enhance the effect of odorant cues for host-seeking mosquitoes [12]; however, our models showed mostly a negative relation, possibly because higher humidity promotes mosquito dispersal [20] and thus decreases the chances of capture. Precipitation on the other hand, significantly increased the chances of capturing blood-fed mosquitoes according to three models; this is counterintuitive as mosquitoes are not expected to fly under rainy conditions. However, this variable was aggregated weekly over the sampling units; thus, higher values do not imply more constant rain but more rain over a week period. Therefore, rainfall may reduce mosquito dispersal increasing the capture chances but does not prevent their feeding activity. Both, Brugman et al. [32] and Karki et al., [45] reported that the wind diminishes the capture of blood-fed mosquitoes, which can be explained by mosquitoes taking shelter under windy conditions, thus reducing the host-seeking activity and the dispersal after blood-feeding. Wind speed was significant in two models but in opposite directions, so we cannot conclude its influence in our samplings. In one model, scarce vegetation was associated with an increase in blood-fed mosquitoes capture suggesting that mosquitoes that are looking for shelter are attracted to the traps in absence of natural resting areas. Finally, a close distance to zoo animal exhibits significantly increased the likelihood of capturing blood-fed mosquitoes in one model, possibly due to dispersal behaviour as discussed before in relation to travelling distances.
After blood feeding, mosquitoes look for a resting place to digest the blood and produce eggs; thus blood-engorged females tend to be captured in low proportions in mosquito traps that target host seeking individuals (e.g. [26]) or gravid females looking to oviposit, which varies depending on the sampling methods and study sites (e.g. [2–4, 22]). It is not clear if blood-fed mosquitoes were attracted by the lactic acid of host-seeking mosquito traps because they are looking for a second blood-meal, or by the fermenting media of gravid traps because they prefer to rest near potential oviposition sites. Alternatively, they may be attracted by the dark colour and location of the traps. However, we cannot exclude the possibility that they were randomly captured as no significant differences were found between BG-Mosquitaire and CDC-Gravid trap types. We captured higher proportions of blood-fed mosquitoes using resting traps and aspirating resting areas, as reported by other authors [2, 32, 48]. In some cases, the use of resting traps has failed [2, 49], thus their positioning and orientation could be a determinant and should ensure that mosquitoes get protection from direct sunlight, rain, and wind.
Our blood-meal identification success could have been influenced by the time before collecting the mosquitoes and weather conditions. If the mosquitoes remained alive, they continued digesting the blood, and if dead, they started to desiccate. We observed more mosquitoes completely dehydrated, despite their evident blood-fed status, during the 2018 sampling in Chester Zoo which was drier and hotter compared to other sampling years. To improve the results, storing the samples at -80 °C or in filter paper and processing them promptly has been recommended [29, 31, 32], which would also minimise physical damage facilitating mosquito morphological identification.
The high proportion of Cx. pipiens mosquitoes feeding on humans that we observed represents not only a likely nuisance for visitors and staff at the zoos, but also a potential risk for disease transmission. Culex spp. are vectors of viruses hosted by wild birds, such as West Nile virus (WNV), Sindbis virus (SINV) and Usutu virus (USUV), all of which have been reported circulating in mainland Europe [17, 50] and could pose a serious threat if they are introduced to the UK. Moreover, all mixed blood-meals that we found were from Cx. pipiens involving a bird host and the mixed blood meals that included humans were combined with blood of Eurasian magpie (Pica pica), which is a proven natural reservoir of WNV and an effective target for its surveillance [51]. In addition, it has been shown that the temporal and spatial variation in host preferences by Culex spp. could influence the timing and severity of WNV infections, probably in relation to its seasonal shifts between ornithophilic and anthropophilic cycles [7, 21]. Although the host shift in Cx. pipiens that we observed in one of our samplings occurred from humans to birds, the potential of this mosquito as a bridge vector for humans and domestic animals (i.e. horses) [49, 52] should be constantly monitored, despite the lack of evidence confirming WNV’s establishment in the UK [17, 53], as well as for the other mentioned viruses (SINV and USUV).
The interspecific transmission of vector borne diseases is also important for the health of the animals in the zoo collections. For instance, mosquitoes have been involved in the transmission of Eastern equine encephalitis virus to African penguins (Spheniscus demersus) in North America, USUV to great grey owls (Strix nebulosa) in Austria and WNV that has caused the death of exotic animals in roughly one hundred zoos in the United States [1]. The risk of disease transmission between local bird species and zoo animals is present in our study sites, as avian malaria has caused outbreaks in Humboldt penguin (Spheniscus humboldti) colonies in both Chester Zoo and Flamingo Land [10]. Interestingly, host preference changes could also influence the transmission dynamics of avian malaria parasites in bird communities [44]. We found four mosquitoes feeding on Humboldt penguins, two Anopheles maculipennis s. l., one Cx. pipiens and an unknown Culicinae, which is likely to be Cx. pipiens as it had all the corresponding morphological features except for those evaluated on the legs. Anopheles spp. mosquitoes are considered as potential vectors for avian Plasmodium spp. and have been found susceptible to the parasite infection experimentally [54]. Therefore, this genus could have a relevant role in the transmission of avian malaria, although An. maculipennis s. l. has not been found infected with avian Plasmodium yet [55, 56]. To clarify the host preferences of this mosquito genus and Plasmodium spp. transmission risks, the precise identification of its species is needed.
{kind=link}
{kind=link}
{kind=link}
{kind=link}