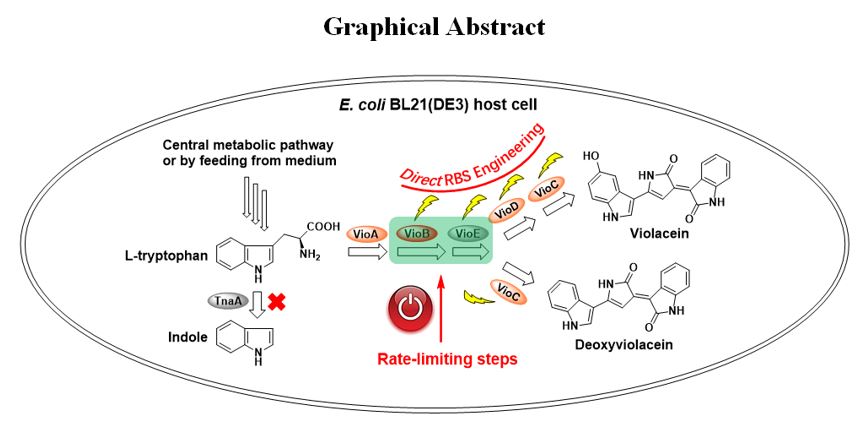
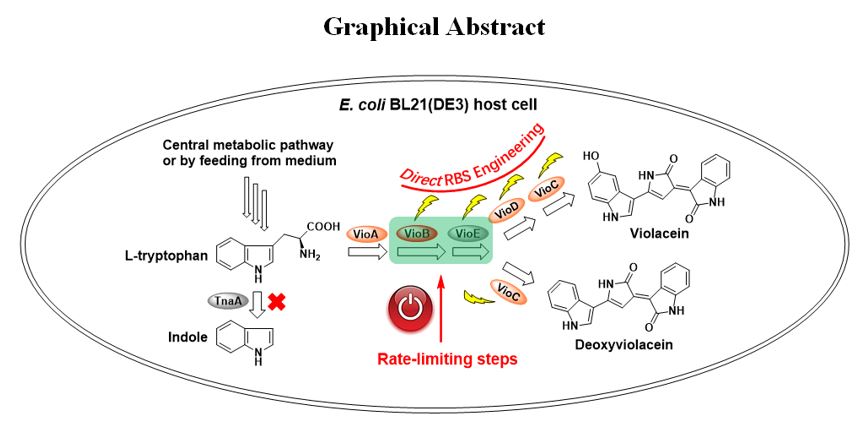
Cloning and heterologous expression of the violaceins biosynthetic gene cluster. Through analyzing the genome sequence of C. violaceum ATCC 12472 by antismash 5.0 software online [29], the five violaceins biosynthetic genes vioABCDE together with its upstream and downstream sequence were obtained. Previously, the upstream sequence of vioABCDE was reported to contain both its natural promoter region and the quorum sensing regulatory site for N-acyl homoserine lactones (AHL) binding [30]. To exclude the possible effect of AHL on this work, we only cloned the five open reading frame vioABCDE, under the control of IPTG induced T7 promoter (Fig. S1). After the generated plasmid Vio12472 was transformed into E. coli BL21(DE3) competent cell, the recombinant strain E. coli BL21(DE3) (Vio12472) had violaceins producing ability (Fig. S2). Strangely, even though IPTG inducer was not added in, light purple bacterial colonies were formed on the solid plate of LB medium at 25 ℃ overnight. This phenomenon may arise from the sporadic gene transcription under the control of T7 promoter (T7 RNA polymerase/lac operon system) induced by small amounts of lactose in the medium which was very complex [31]. The colonies’ color turned into deep purple when 0.1 mM IPTG was used in the medium (Fig. S2A). To determine the metabolite produced by this recombinant strain, the crude violaceins were analyzed by liquid chromatograph-mass spectrum (LC-MS) system. The results showed that two species, violacein and deoxyviolacein, were produced at the same time (Fig. S3).
Four rounds of direct RBS engineering of the violacein biosynthetic gene cluster.
In this work, vioABCDE operon was under the control of a strong T7 promotor but the five RBSs were apparently different (“AAGGAG” for vioA, “GGGAAA” for vioB, “GAGAGG” for vioC, “AGGGAG” for vioD and “AGGAGG” for vioE) (Fig. 2A). In addition, RBS of vioC was located within the ORF of vioB while vioC and vioD formed as overlapping genes. Because RBS of vioA gene was a strong RBS in E. coli host strain, we supposed that engineering the native RBSs of other four genes (vioB, vioC, vioD and vioE) may further improve corresponding translational efficiency to enhance the final production of violaceins. Therefore, this work carried out four rounds of direct RBS engineering of violaceins biosynthetic gene cluster by inverse PCR technique to introduce in-situ site-specific mutagenesis, including DNA base-pairs exchange and deletion (Fig. 2B). The cultivation of wild type strains and the mutants were all carried out in flask with 50 ml LB broth and the initial fermentation conditions were the same as follows: After incubating to OD600 nm= 0.8 at 37 ℃, the temperature was shifted to 25 ℃ and 0.1 mM IPTG inducer was simultaneously added for another 24 h’s fermentation.
In the first round of RBS engineering, we respectively introduced a single RBS mutation to the violaceins biosynthetic gene cluster, yielding seven mutated strains named as Bm-1, Bm-2, Bm-3, Cm, Dm-1, Dm-2 and Em (Fig. S5). Because VioB protein was verified as one rate-limiting enzyme in the biosynthetic pathway of violaciens in the previous report [21], we firstly constructed three RBS mutants of vioB gene. Bm-1 and Bm-2 both contained partial sequence (“GGGGAG” for Bm-1 and “AAGAAA” for Bm-2) of the strong artificial RBS (AAGGAG) while Bm-3 had the same RBS as vioA gene (Fig. S5). Compared with the starting strain E. coli BL21(DE3) (Vio12472), RBS mutagenesis of vioB apparently affected the total yields of violaceins (Fig. 3A). The titers of Bm-2 and Bm-3 were respectively 1.17 and 1.25 times to the wild type while Bm-1’s titers were reduced to 90%. These results suggested the strong RBS (AAGGAG) was more effective than vioB’s native RBS (GGGAAA). Next, we constructed the RBS mutant of vioE gene which was also verified to catalyze another rate-limiting step in the violaceins biosynthesis [22] (Fig. S5). After the native RBS (AGGAGG) of vioE was mutated to RBS (AAGGAG), the productivity of Em was 1.01 times to the starting E. coli BL21(DE3) (Vio12472) (Fig. 3A). Because the native RBSs of vioC and vioD were respectively overlapped with the ORFs of vioB and vioC, this study didn’t carry out site-specific mutagenesis to prevent introducing frame shift mutation to the upstream genes. For this case, we adopted DNA base-pairs deletion strategy to shorten the distance of RBS with its downstream initiation codon “ATG”, evaluating the effect of this type of mutagenesis on the production of violaceins. Deletion of “AA” base-pairs between the ORF of vioB and vioC gene was firstly carried out to obtain a new vioBC overlapping gene (the termination codon TGA of vioB shared the same “A” with the initiation codon “ATG” of vioC in the mutant Cm) (Fig. 2A and Fig. S5). Cm could produce 1.94 times of total violaceins to the wild type, especially 2.78 times of deoxyviolacein component (Fig. 3A). This result suggested the newly formed vioBC overlapping gene was helpful to VioC protein’s translation and then acted on the metabolic flux to produce more deoxyviolacein. For RBS engineering of the vioD gene, Dm-1 was firstly constructed by deletion of “GTC” codon from the 3’-terminal of vioC gene (note: the “GTC” codon encodes the second amino acid from the C-terminal, which was not locating within the active site of VioC protein and therefore its deletion should not affect the enzymatic activity or stability of this enzyme) (Fig. S5). The fermentation titers of Dm-1 were 1.16 times to the wild type (Fig. 3A). Based on Dm-1, we further deleted another three base-pairs “AAC” (downstream of “GTC” codon) to construct a new mutant strain Dm-2 (Fig. S5). However, the production of violacein by Dm-2 was only 37% of the wild type while deoxyviolacein’s yield was 3.52 times to that of the wild type strain (Fig. 3A). These results indicated that the mutation in Dm-2 probably reduced the translation level of VioD protein. The direct RBS mutagenesis didn’t change the growth situation because obvious differences of the biomass between the wild type and the mutants were not found. They were all about 1.5 g CDW L− 1 (Fig. 3B).
Through the first round-engineering on the native RBS of violaceins biosynthetic gene cluster, five higher producing mutant strains Bm-2, Bm-3, Cm, Dm-1 and Em were obtained. Next, this study wanted to know whether combinational RBS mutagenesis of two genes at the same time could further enhance the production titers of violaceins, so another six mutant strains were constructed, including (BC)m, (BD)m, (BE)m, (CD)m, (CE)m, (DE)m. In the cases of (BC)m, (BD)m, (BE)m, we adopted the RBS mutation type of Bm-3 in the first round-mutagenesis which was verified as the best mutant. By batch fermentation in flasks, the yields of these six mutants were quantified by HPLC (Fig. 3C). The results showed combinational RBS mutations had apparently positive effect on the production of violaceins, especially the (BE)m strain whose total yields of violaceins were 2.21-times to the wild type. Furthermore, compared with the starting strain E. coli BL21(DE3) (Vio12472) and two single RBS mutants (Bm-3 and Em), the color of (BE)m on the solid plate (without IPTG inducer used) was obviously deeper purple (Fig. 4A). Strangely, the overexpression of VioB and VioE proteins in (BE)m was not detected by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) (Fig. S6). We predicted that the combinational RBS engineering of vioB and vioE, catalyzing two known verified rate-limiting steps in the biosynthetic assembly line of violaceins [21–22], may probably break through the bottleneck of intermediate conversion. There were no obvious differences of the biomass between the wild type and the mutants (Fig. 3D).
Like the second round-engineering on the native RBSs of violaciens biosynthetic gene cluster, we further carried out combinational RBS mutagenesis of three genes at the same time, generating four mutant strains including (BCD)m, (BCE)m, (BDE)m, (CDE)m (Fig. 5A). The (BCD)m and (BCE)m were constructed based on (BC)m while (BDE)m based on (BD)m and (CDE)m based on (CD)m. The product yields of these four mutants also revealed the positive effect of combinational RBS mutations on the production of violaciens. The total violaciens yields of (BCD)m, (BCE)m, (BDE)m, (CDE)m were respectively 1.60, 2.16, 2.01, 1.39 times to the wild type E. coli BL21(DE3) (Vio12472). On the solid plate of LB medium, the colony color of (BCE)m and (BDE)m were also deep purple like (BE)m (Fig. 4A), once again suggesting that when the native RBSs of vioB and vioE were both mutated to the strong RBS (AAGGAG), the metabolic flux of violaceins biosynthesis may become smoother with less of obstruction.
Finally, this work also constructed a mutant strain (BCDE)m based on (CDE)m by combinational RBS mutagenesis of four genes (vioB, vioC, vioD and vioE) (Fig. 4B). The color of (BCDE)m on the solid plate was also deep purple, same as other three mutant strains, (BE)m, (BCE)m and (BDE)m (Fig. 4). The fermentation yield of violaceins by (BCDE)m was 2.41-fold improvement to the wild type strain E. coli BL21(DE3) (Vio12472) and 1.73- fold improvement to its starting strain (CDE)m. However, a comparison of the titers of (BCDE)m, (BCE)m, and (BDE)m strains with that of (BE)m strain suggested RBS engineering of vioC and vioD genes could not make a significant contribution on the violaceins biosynthesis. Based on these results, we deduced that in the biosynthetic routes of those RBS engineered violaceins biosynthetic gene cluster, the oxidation steps catalyzed by VioC and VioD may become “newly” rate-limiting steps. The direct RBS engineering strategy used in this work may break through the natural balance of five proteins and create new balance of them in the mutant violaceins biosynthetic gene clusters. In the future, the engineering of the steps catalyzed by VioC and VioD proteins with new method may further improve the titers of violaceins. The multiple combinational RBS mutagenesis didn’t have negative effect on the biomass (Fig. S7A).
Optimization of the cultivation conditions of the RBS-engineered highest producer (BCDE)m.
After obtaining the highest producing strain (BCDE)m, this work further intended to determine its productivity of violaceins at the optimal conditions including temperature, concentration of IPTG inducer and fermentation time. One-factor-at-a-time method was used in this work. Firstly, the strain (BCDE)m of five groups were cultivated in 250 ml shake flask with 50 ml LB medium broth at 37 ℃ to OD600 nm about 0.8. and then temperatures were separately changed to 16 ℃, 20 ℃, 25 ℃, 30 ℃ and 37 ℃ together with adding 0.1 mM IPTG inducer for another 24 h’s fermentation (Fig. 5B). The results showed that at 30 ℃ the productivity of (BCDE)m was highest and the total violaceins yields reached to 1717.6 µM. Strangely, we found temperature at 37 ℃ was also available for the efficient production of total violaciens by (BCDE)m with yields of 1445.0 µM and even higher titers for the violacein component than that at 30 ℃. For this case, we speculated that higher temperature such as 37 ℃ may be good for the activity of VioD protein which was a key enzyme for regulating the ratio of violacein and deoxyviolacein components in the total products. These liquid fermentation results also supported the phenomenon that (BCDE)m formed apparent deep purple colony on the solid plate after overnight cultivation (about 12–14 h) at 37 ℃ even without IPTG inducer. In the previous reports, all of the violaciens producing strains including the natural producers and recombinant producers could only work well at lower temperatures and 20 ℃ was mainly recognized as the most optimal condition for the fermentation of violaceins because elevated temperature may destroy the protein folding of violaciens biosynthetic enzymes, leading to remarkable reduced productivity [13, 15]. One report even showed that Pseudoalteromonas sp. 520P1, a natural violaceins producer, would not survive at 37 ℃ [30]. Therefore, it is the first time to find that RBS engineered violaceins recombinant producing strains could work efficiently at higher temperatures. In consideration of the apparent phenotypes on the solid plate between the series of (BE)m mutants and their basic strain E. coli BL21(DE3) (Vio12472), we speculated that the combinational replacing of the natural RBSs of two rate-limiting enzyme encoding genes, vioB and vioE, with stronger artificial RBS (AAGGAG) may break through the metabolic bottleneck. To our knowledge, fermentation at higher temperatures (such as 30 ℃ and 37 ℃) has a lot of advantages than lower temperatures (such as 20 ℃ and 25 ℃), energy loss caused by cooling measures could be remarkably reduced. For this case, we will further explore the reasons for this interesting phenomenon in the future. Because the biomass at 37 ℃ was a little more than 30 ℃ (Fig. S7B) while its total productivity was lower than 30 ℃ (Fig. 5B), we decided 30 ℃ as the basis for further optimizing.
Other than temperature, IPTG also plays an important role in heterologous protein expression and affecting the productivity of the host strain. However, excessive IPTG not only had toxicity to the normal growth of cell,31 but also would accelerate the expression speed of target proteins to result in the formation of a large portion of inclusion bodies [32]. Therefore, to find an optimal value of IPTG inducer, this study tested various concentrations of IPTG including 0.01, 0.02, 0.04, 0.06, 0.08, 0.10, 0.20, 0.40, 0.60, 0.80 and 1.00 mM (Fig. 5C). The fermentation results showed that 0.02 mM IPTG was the best amount for violaceins production by (BCDE)m with a total titer of 2095.9 µM while all the IPTG concentrations used in this work didn’t apparently affect the biomass (Fig. S7C). These data showed that at the higher concentration of IPTG (over 0.06 mM), the production of violacein component became reduced while deoxyviolacein component increased apparently. In addition, the productions of total violaceins at higher concentration of IPTG were also lower (Fig. 5C). For this phenomenon, we deduced that higher usage of IPTG may negatively affect the expression level or the catalytic activity of VioD and VioC protein which were two final steps for the formation of violaceins. As our speculation above, the balance of the five proteins in the RBS engineered violaceins biosynthetic gene cluster may be different from the wild type, the oxidation steps catalyzed by VioC and VioD could become “newly” rate-limiting steps.
Next, based on the optimized temperature and IPTG concentration, we carried out the fermentation time test for (BCDE)m strain because appropriate fermentation period was very important for future practical application, especially at the industrial scale. Long fermentation time would not only increase the cost of production including more raw material consumption and more administration expense, but also affect the product quality because a lot of fermentation products were not stable for a long time in the complicated physicochemical environment (including the changes of pH, oxygen concentration, temperature, light strength) of fermentation broth. In addition, longer fermentation time may increase the possibility of contamination by other microorganisms from the external environment. Therefore, this study optimized the cultivation period of (BCDE)m strain by batch fermentation from 24 h to 192 h (Fig. 5D). These results showed that the total yield of violaceins at 48 h (2382.6 µM) reached the highest. With the fermentation time extending, the productivity of (BCDE)m decreased gradually. The biomass also followed the similar trend to violaceins’ yields (Fig. S7D). We speculated that the problems should come from nutrient deficiency and cell death at the late-stage of fermentation. Finally, based on the series of experiment above, the optimal condition for the fermentation of (BCDE)m strain was summarized as follows: After cultivating to OD600 nm = 0.8 at 37 ℃, the temperature was turned down to 30 ℃ and at the same time 0.02 mM IPTG inducer was added in the LB medium for another 48 h’s fermentation.
The productivity of (BCDE)m further improved by feeding tryptophan.
After obtaining the optimal fermentation conditions of the mutant strain (BCDE)m, this work further intended to test its productive potential through feeding with excess amount of the precursor L-tryptophan. In the metabolic network of tryptophan, tnaA gene encodes tryptophanase which is responsible for degradation of intracellular free tryptophan into indole. Indole molecules had toxicity to the cell growth at high concentration [33]. To exclude possible negative effects of tnaA gene on the production of violaciens in this study, we constructed a tnaA-knocking out strain of E. coli BL21(DE3) by λ-red homologous recombinant method (Fig. S8). Then, the plasmid Vio12472-vioBCDE-RBSm and its wild type plasmid Vio12472 were respectively transformed into the E. coli BL21(DE3) (tanA−) to generate two new strains (BCDE)m (tanA−) and Vio12472 (tanA−).
Finally, under the optimal fermentation conditions, we carried out batch fermentations for the two new strains by adding various concentrations of tryptophan (0, 1, 2, 4, 6, 8, 10 mM) to the LB medium together with IPTG. After cultivation of 48 h, the yields of vioalceins of (BCDE)m (tanA−) reached summit (3269.7 µM) when feeding with 2 mM tryptophan while the control strain Vio12472 (tanA−) also reached the highest production (2284.0 µM), about 69.9% of the (BCDE)m (tanA−) (Fig. 6A and 6B). However, the apparent improvement of yields along with the increased usage of L-tryptophan for both (BCDE)m (tanA−) and Vio12472 (tanA−) did not appear. On the contrary, excess tryptophan (above 4 mM) negatively affected the productivity of violaceins. We measured the tryptophan consumption and found there were large amounts of residues at higher concentrations of this precursor in the medium (Fig. 6C, 6D and S4). Therefore, we speculated that violaceins may accumulate upon the cell surface and block the entrance of tryptophan precursor into the cell because of the poor water solubility of violaceins in the medium. To further improve the productivity of the RBS engineered violaciens biosynthetic gene cluster in the future study, we plan to do the in vivo experiments based on the tryptophan hyperproducer, such as tryptophan biosynthetic pathway well optimized E. coli [13] or C. glutamicum host strains [15].
{kind=link}
{kind=link}