G. hirsutum genome contains 81 GhWAKs
The known A. thaliana WAK protein sequences were used as queries to search the genome database of G. hirsutum accession Texas Marker-1 (TM-1). In total, 81 GhWAKs as candidates were identified and named according to their chromosomal locations. These GhWAKs were marked on the physical map of 18 chromosomes (Figure 1) and one scaffold664 (GhWAK65). A total of 34 and 46 GhWAKs were distributed in the A and D sub-genomes, respectively. Chromosome D02 harbored the largest number of GhWAKs with 20 genes. Six pairs of tandem duplication events were found, including GhWAK16/17, GhWAK36/37, GhWAK43/44-49, GhWAK50/52, GhWAK61/62 and GhWAK69/70/71. These results revealed that the evolution and expansion of GhWAKs happened in G. hirsutum, especially on chromosome D02. The detailed information about GhWAKs, including gene ID, open reading frame length, amino acid length, protein molecular weight and isoelectric point, instability index and subcellular localization, was listed in Table 1.
All GhWAKs were predicated to be localized on plasma membrane (PM) (Table 1). In which, GhWAK77, as a representative, its location was further confirmed using transient expression of fluorescent fusion proteins in tobacco epidermal cells. The images clearly showed that fluorescent signal corresponding to the sole gfp gene was observed in PM, cytoplasm and nucleus. However, the fluorescent signal corresponding to GhWAK77-gfp was solely shown in PM (Figure 2). These suggest that GhWAKs have potential as connector responsible for communication between inside and outside of the cell.
GhWAKs haveconservative kinase domains and various extracellular domains
The majority of GhWAKs have 3-4 introns and show similar exon-intron structure (Figure 3A). GhWAK36 contains the maximum number introns (six), whereas GhWAK11 and GhWAK53 do not contain any intron. A total of six conserved protein domains were identified in GhWAKs, including GUB_WAK_bind (wall-associated receptor kinase galacturonan-binding, PF13947), WAK (wall-associated kinase, PF08488), WAK assoc (wall-associated receptor kinase C-terminal, PF14380), EGF (EGF, PF00008; cEGF, PF12662; hEGF, PF12661; EGF_CA, PF07645; EGF_3, PF12947), DUF1199 (domain of unknown function, PF06712) and protein kinase domain (pkinase, PF00069; pkinase_Tyr, PF07714; kinase-like, PF14531; protein-kinase domain of FAM69, PF12260) (Figure 3B). Cytoplasmic, extracellular and TM regions were predicated in the majority of GhWAKs, further indicating that they are membrane proteins. Typical WAK encodes a transmembrane protein with a cytoplasmic kinase domain and an extracellular region. However, several proteins showed uncommon structural characteristics, such as the kinase domain in extracellular region (GhWAK43, GhWAK54, GhWAK75, GhWAK57, GhWAK2, GhWAK36, GhWAK37 and GhWAK40), double TMs (GhWAK18, GhWAK58, GhWAK59, GhWAK60, GhWAK64, GhWAK75 and GhWAK1) and kinase domains (GhWAK59). All GhWAKs were classified into seven groups according to their protein domain analysis (Figure 3B and 3C). The members in Group I, Group II and Group III were typical WAKs that contain EGF domain in extracellular region. The other four Groups, including IV, V, VI and VII, do not contain EGF. GhWAKs in Group I and IV contain both WAK and GUB domain. Inversely, GhWAKs in Group III neither contain WAK nor GUB domain. GhWAKs in Group II, VI and VII only contain GUB domain. However, II and VI are one-GUB-domain groups, and VII are two-GUB-domain group. GhWAKs in Group V only contain WAK domain. In additionally, DUF1199 domain was found in GhWAK31 and GhWAK77 (Figure 3B). Different types and numbers of extracellular domains were present in GhWAKs, indicating that they might sense or bind different outside signaling to trigger special intracellular signaling pathways that control plant development and responses to various environmental stresses.
Prediction of putative cis-regulatory elements in GhWAK promoters
The 2 kb region upstream of the translation start site of all GhWAKs were considered the promoter and analyzed for investigating the potential roles of cis-regulatory elements. The numbers and names of identified cis-elements were shown on Figure 4. These cis-regulatory elements were classified into four main groups including hormones, biotic, abiotic and light. Twelve hormone-responsive regulatory elements associated with abscisic acid (ABA) (ABRE, ABRE4 and AT-ABRE), auxin (IAA) (AuxRR-core, TGA-box and TGA-element), methyl jasmonate (MeJA) (CGTCA-motif), gibberellin (GA) (GARE-motif, P-box and TATC-box), SA (TCA-element) and ethylene (ET) (ERE), were identified. Of which, ABRE-motif, CGTCA-motif and ERE were enriched in the most of GhWAK promoters, indicating that they might be widely induced by ABA, JA and ET. The biotic stress-related regulatory elements, such as AT-rich, TC-rich repeats, W-box, WUN-motif, WRE3, JERE and box S, were involved in elicitor-mediated activation, wounding and pathogen responsiveness. In additionally, eight abiotic-responsive regulatory elements, associated with anaerobic induction (ARE and GC-motif), low-temperature responsiveness (LTR), drought-inducibility (MBS, DRE core and DRE1), heat shock, osmotic stress, low pH, nutrient starvation (STRE) and stress-related (TCA), were identified in the GhWAK promoter regions. Moreover, various light-responsive elements were present in the promoters of GhWAKs. Especially, Box 4 and G-Box were widely harbored. These results indicated that GhWAKs might play vital roles in the response to various stresses, hormones and light.
GhWAKs were significantly induced by V. dahliae infection
To identify GhWAKs that are related to V. dahliae infection, two-fold changes were applied in transcript expression profiles from RNA-seq as minimum cutoffs. As a result, 26 GhWAKs were screened out, including 17 up-regulated and 9 down-regulated genes (Figure 5). Of these, 11 GhWAKs, including GhWAK5, GhWAK9, GhWAK77, GhWAK10, GhWAK45, GhWAK47, GhWAK78, GhWAK48, GhWAK31, GhWAK26 and GhWAK72, were significantly up-regulated in at least three time points. Further, the expression profiles of GhWAKs in response to the infection with V. dahliae were detected and verified through qRT-PCR (Figure 6). GhWAK1 and GhWAK69 showing higher transcription levels in roots of cotton seedlings inoculated with V. dahliae than control was screened out. Due to the high degree of sequence similarity in GhWAKs family, it is difficult to design specific primers for four gene pairs, including GhWAK4 & GhWAK45, GhWAK5 & GhWAK49, GhWAK10 &GhWAK55, and GhWAK31 & GhWAK77. The results of qRT-PCR indicated that these four pairs of genes were dramatically up-regulated. According to RNA-seq data, GhWAK4, GhWAK49 and GhWAK55 did not shown to be up-regulated. Thus, the expression changes found using qRT-PCR probably more represent the responses of GhWAK45, GhWAK5 and GhWAK10 to V. dahliae infection. The expression results of GhWAK26, GhWAK48 and GhWAK72 were consistent with RNA-seq data with up-regulation. In total, 31 GhWAKs were screened out, which play a potential role in the interaction between cotton and V. dahliae.
Silencing of GhWAKscompromises cotton resistance to Verticillium wilt
GhWAK26 and GhWAK77 showed obviously persistent up-regulated expression to the infection from V. dahliae (Figure 5 and Figure 6). In addition, they contain cis-elements in their promoters associated with MeJA and SA, which play key roles in cotton resistance to V. dahliae. Thus, to further reveal the function of GhWAKs in cotton resistance to V. dahliae, GhWAK26 and GhWAK77 were prioritized for study as representatives in this study using TRV based VIGS system. At approximately two weeks post-infiltration with a mixture of Agrobacterium cultures containing pTRV1 and pTRV2-CLA1, a strong photobleaching phenotype was shown on the newly emerging true leaves (Figure 7A), indicating that VIGS system worked well. Then, the expression levels of GhWAK26 and GhWAK77 were detected in the leaves infiltrated with pTRV2-GhWAK26 and pTRV2-GhWAK77, respectively. As shown in Figure 7B, the expression levels of GhWAK26 and GhWAK77 were reduced by about 80%, suggesting VIGS triggered their silencing in cotton plants. At 20 dpi, VIGSed plants (Figure 7D and 7E) exhibited more chlorosis and wilting than the control plants infiltrated with Agrobacterium cultures containing empty vector pTRV1 and pTRV2 (Figure 7C). The disease indices of VIGSed plants were also significantly higher than those of the control at 15 dpi and 20 dpi (Figure 7F). Therefore, the results of VIGS assays suggested that GhWAK26 and GhWAK77 are important participants in cotton resistance to V. dahliae infection.
Silencing of GhWAKs significantly affected the expression of JA and SA-related marker genes
Further, the expression of several JA and SA-related marker genes involved in plant defense signaling pathways was detected. The expression of JAZ1 (jasmonate-zim-domain protein), JAZ3, JAZ6, LOX1 (lipoxygenase) (JA-related marker genes), PR3 (pathogenesis related protein) and NPR1 (nonexpresser of PR protein) (SA-related marker genes) were significantly down-regulated after silencing of GhWAK26 in cotton (Figure 8A). In GhWAK77-silenced plants, JAZ6 and three important genes involved in the SA signaling pathway, including ICS1 (isochorismate synthase), NPR1 and EDS1 (enhanced disease susceptibility), were down-regulated comparing with control. On the contrary, the expression of JAZ1 and LOX1 were significantly up-regulated due to the silencing of GhWAK77 (Figure 8B). These indicated that GhWAK26 and GhWAK77 may involve in cotton resistance to V. dahliae by SA and JA signaling passway.
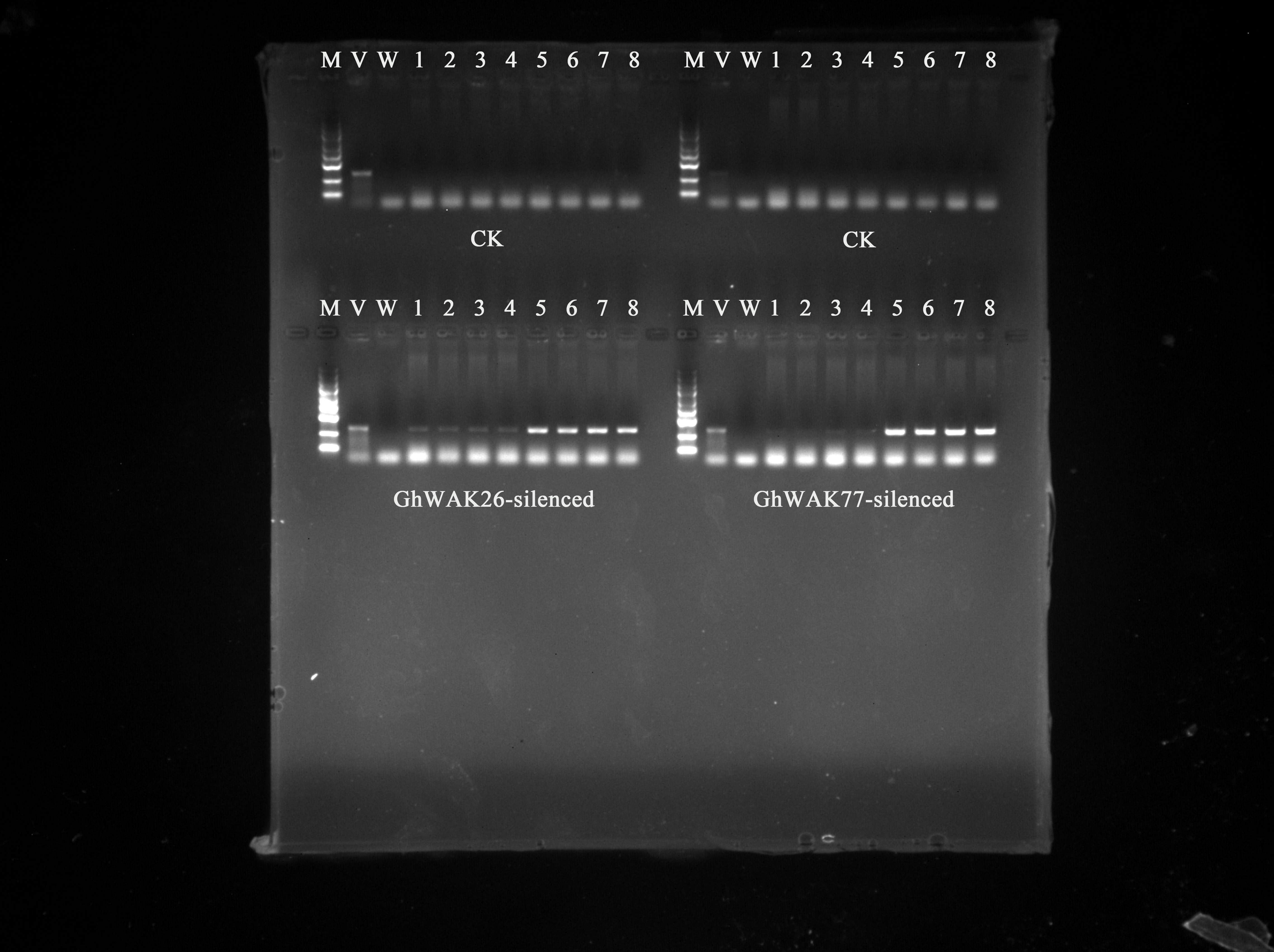
Silencing of GhWAKs increased the spread of V. dahliae in the cotton stems
After inoculation, V. dahliae in cotton stems was detected by PCR. As shown in Figure 9A, no specific amplification products from V. dahliae were shown in CK at 5 dpi and 7 dpi, indicating that V. dahliae had not yet invaded the stems or multiplied in large quantities. However, at 5 dpi, few specific products from V. dahliae were amplified in GhWAK26-silenced and GhWAK77-silenced plants, representing a small amount of pathogen invasion. Further, at 7 dpi, the bright bands amplified using stems from GhWAK26-silenced and GhWAK77-silenced plants appeared on agarose gels, indicating that V. dahliae had invaded largely. In addition, pathogen isolation on PDA showed that a large number of V. dahliae grew out from the stems of GhWAK26-silenced and GhWAK77-silenced cotton plants, while no mycelium was shown from the control (Figure 9B). Both PCR detection and PDA culture results suggested that silencing GhWAKs significantly increased the spread of V. dahliae in the cotton stems.
Lignin is considered to play an important role in preventing the infection of V. dahliae in cotton. Therefore, we further compared the changes of lignin content in GhWAK-silenced cotton stems with CK. The results showed that the lignin content in GhWAK-silenced plants was significantly lower than that in CK (Figure 9C), which might affect the stem structure and then reduce the prevention of cotton on V. dahliae infection.
Silencing of GhWAKs dramatically compromised V. dahliae-induced accumulation of H2O2 and NO (nitric oxide), but enhanced POD (peroxidase) activity
The contents of H2O2 and NO, and POD activity in GhWAK-silenced plants inoculated with V. dahliae were further measured. GhWAKs silencing caused lower levels of H2O2 at 6 hpi (hours post inoculation), 12 hpi and 24 hpi (Figure 10A and 10B). Both GhWAK26- and GhWAK77-silenced plants accumulated greatly depressed levels of NO comparing with CK (Figure 10C and 10D). However, the activity of POD significantly elevated in GhWAK26- and GhWAK77-silenced plants at 6 hpi, 24 hpi and 48 hpi, except at 12 hpi (Figure 10E and 10F).
{kind=link}