Quality assessment of the anti-T. cruzi and cells toxicity assays.
As part of the process of setting up the biological assays we calculated their Z´ parameter to assess reproducibility and statistical robustness (23). In general, assays with a Z´ between 0.5 and 1 are considered appropriate for the screening of compounds (23). Remarkably, our anti-parasitic assay had a very good performance and its Z´ remained consistently > 0.5 with an average value of 0.89 (0.097) (Figure 2A). Regarding the two cytotoxicity assays used in this work, we respectively retrieved Z´ = 0.76 (0.067) for the assay based on Vero cells, and a Z´ = 0.73 (0.054) for the assay based on HepG2 cells (Figure 2C-E).
Additionally, in every run of the T. cruzi growth inhibition assay and Vero cells toxicity assays performed, we included the reference drug BNZ as control, whereas the reference drug digitoxin (DTX) (24) was included in all the HepG2 cells toxicity assays. Overall, averaged IC50 and TC50 values for BNZ were respectively 1.56 (0.39) μM and 173.4 (43.57) μM (Figure 2B-D; Figure S3), which correlate with previous reports (21, 25). Digitoxin TC50 mean value in the HepG2 cell assay was 0.29 (0.14) μM (Figure 2F; Figure S3).
Figure 2. Quality controls of the T. cruzi growth inhibition assay (A-B), toxicity assay with Vero cells (C-D) and toxicity assay with HepG2 cells (E-F). Z´ values for each of the rounds launched are represented on the left (A, C, E); dashed line marks the 0.5 threshold. IC50 and TC50 values of the reference drugs BNZ and DTX are represented on the right (B, D, F); straight lines indicate the average values, whereas the dashed lines indicate ± 3 SD limits.
Anti-T. cruzi activity of the alkaloids extracted from Amaryllidaceae.
Same as other widely used anti-T. cruzi assays (21, 22, 26) we relied on the genetically robust T. cruzi - Tulahuen strain expressing beta-galactosidase activity as a surrogate of parasite growth (15). However, being the amastigote replicative stage of T. cruzi obliged intracellular, our assay relies on Vero cells as hosts. Notably, Vero cells were described to be deficient in the interferon response due to a mutation in the simian interferon beta gene (27), which makes them more susceptible to infections and thereby a good system for the discovery of active compounds against T. cruzi.
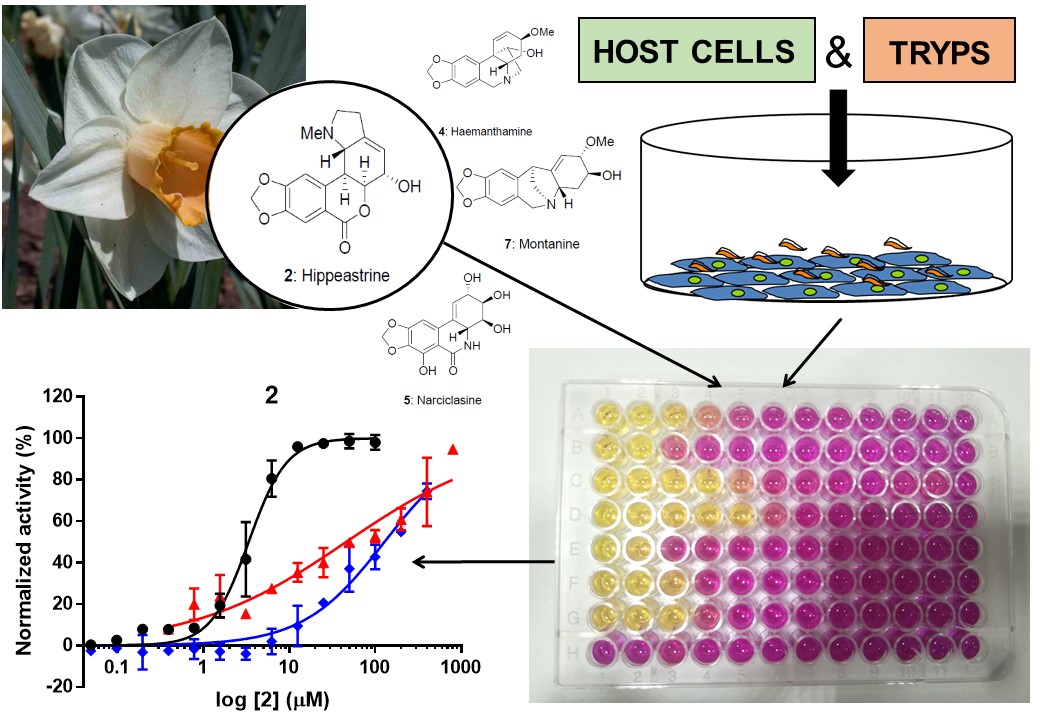
Results obtained from the phenotypic T. cruzi growth inhibition assay revealed that lycorine 1, hippeastrine 2, haemanthamine 4, narciclasine 5 and montanine 7 were active, while crinine 3, tazettine 6, sanguinine 8 and 1-O-acetylcaranine 9 were inactive against the parasite (Figure 3). Tazettine 6, sanguinine 8 and 1-O-acetylcaranine 9 have been described as poor antiprotozoal agents before (28, 29). Nonetheless, Machocho and co-workers described that compound 3-O-acetylsanguinine had some activity (IC50 = 2.3 μg/mL; i.e. 7.29 μM) against trypomastigotes from T. cruzi strain Tulahuen C4 (28). Although different phenotypic anti-T. cruzi assays were performed in each case, the results obtained by those authors could suggest that the presence of an acetyl group might increase the anti-T. cruzi activity of sanguinine.
On the other hand, crinine 3 and haemanthamine 4 are crinane-type alkaloids that belong to the β-crinane and α-crinane subgroups, respectively (30). Other studies evaluating the anti-parasitic potential of alkaloids have reported that the presence of a methylene-dioxi group seemed to favour a more potent anti-parasitic activity (31). This could be the explanation for the different anti-T. cruzi activities observed by us between these two crinane-type alkaloids.
The highest anti-T. cruzi activity rates were yielded by lycorine 1 [IC50 = 0.70 (0.018) µM] and narciclassine 5 [IC50 = 0.495 (0.018) µM], which exceeded in potency that of the reference drug BNZ (Table 1; Figure S3). Lycorine 1 was the first alkaloid described from the Amaryllidaceae plant family and one of the most commonly found amongst different genus (32). Lycorine 1 extracted from Crinumstuhlmannii, Zhepyranthes citrina and Narcissus broussonetii had been tested before against T. cruzi (33–35). However, in contrast to our results, it was described as inactive against the parasite; a feature that could be explained due to the use of different assays. It has been reported that major differences on compounds activity can be found depending on the host cell lines and the biological assays used (36). In this respect previous references relied on the use of L6 cells as hosts, whereas we have used Vero cells. Lycorine 1 and 1-O-acetylcaranine 9 belong to the lycorine group but structurally differ in two positions of the structure (Figure 1). In this case, the anti-parasitic activity is increased with the substitution of a hydrogen and an acetoxy group per two hydroxyl groups.
With respect to narciclasine 5, which yielded the most potent activity against T. cruzi in this study (Table 1), to the best of our knowledge it has not been tested against T. cruzi before. Likewise, there were no previous reports on the anti-T. cruzi activity of montanine 7, also shown for the first time in this study. Montanine 7 and haemanthamine 4 showed averaged IC50 values similar to that of BNZ, respectively 1.99 (0.089) µM and 1.59 (0.062) µM versus 1.56 (0.070) µM of BNZ (Table 1; Figure S3). Osorio and co-workers had previously described the high activity of haemanthamine against T. cruzi (IC50 = 1.8 µg/mL; i.e. 5.97μM) (29).
Finally, we found that hippeastrine 2 was the active alkaloid with the higher IC50 value [IC50 = 3.63 (0.24) µM]. Its activity was poorer than that of BNZ but still fell within a range five times the IC50 of the standard drug (Table 1; Figure S3). Hippeastrine 2 was first isolated from the Amaryllidaceae plant Lycoris radiata and reported to exhibit activity against avian influenza virus H5N1 [IC50 = 47.5 (0.37) µM] (37). Moreover, promising results against ZIKA virus infection have been recently reported with hippeastrine hydrobromide (38). This was shown to remove ZIKA virus presence from infected human neural progenitors, recover a ZIKV-induced microcephaly phenotype in human forebrain organoids and even suppress virus propagation in infected adult mice (38). Antiviral (37, 38), antibacterial and antifungal (39) activities have been reported for hippeastrine 2, despite little information is available about its anti-parasitic activity. Cedron et al. tested twenty one hippeastrine 2 derivatives that included functional group transformations, structural simplification and dimers formation against Plasmodium falciparum (strain F-32 Tanzania) (40). The anti-malarial activity increased by 10-fold when dimers were evaluated compared to the single alkaloid activity, suggesting an improved binding to the related target or the hydrolysis of the dimer onto two molecules (40). To our knowledge this is the first time that anti-T. cruzi activity is reported for hippeastrine 2. Results reported by Cedron and co-workers would suggest to further pursue research with hippeastrine 2 derivatives against T. cruzi.
Identification of alkaloid compounds with specific anti-T. cruzi activity
With the aim of further selecting those alkaloids with specific activity against the parasite and discard those that were toxic to host cells we used two secondary cell toxicity assays with monkey (Vero) and human (HepG2) cells. Since the compounds activity might vary depending on the characteristics of the cell line used, assaying cytotoxicity in two cell lines will provide a more robust readout. Moreover, HepG2 cells are a widespread cell model to anticipate potential liver toxicity of drugs metabolism (41, 42). We determined a selectivity index (SI; or TC50 to IC50 ratio) > 10 to consider an alkaloid for further progression as described elsewhere (22).
Thereafter, all the alkaloids reported as active against T. cruzi were analysed through both cell toxicity assays. All of them were more toxic to Vero cells than to HepG2 cells (Figure 4). The cytotoxicity values registered against Vero and HepG2 cells indicated that narciclasine 5 activity was not specific against T. cruzi [TC50 = 0.66 (0.082) µM against Vero cells; and TC50 = 2.73 (0.67) µM against HepG2 cells, resulting in SI < 10 in both cases (Table 1). In addition, montanine 7 with a SI = 2.5 in relation to Vero cells was not specific to T. cruzi either (Table 1), even though it showed low toxicity to HepG2 cells [TC50 = 46.1 (11.99) µM] (Table 1). Something similar occurred with lycorine 1, which was weakly active against HepG2 cells [TC50 = 21.87 (4.16) µM] (Table 1), presenting a SI versus this cell line over ten times its registered anti-T. cruzi activity, but turned to be toxic to Vero cells [TC50 = 5.21 (0.80) µM] (Table 1) yielding a SI < 10 and thus being discarded it from further progression. We generally observed an increased sensitivity of Vero cells to the alkaloids when compared to HepG2 cells. This may have been in part due to the fact that compounds incubated for longer time (four versus two days) on Vero cells than on HepG2 cells.
Haemanthamine 4 had been reported to present a TC50 = 13 µg/mL; i.e. 43.14 µM for HepG2 cells (43) which correlate with the TC50 value we have retrieved in our study with this cell line [TC50 = 42.48 (6.96) µM] (Table 1). But again, although it showed a SI > 10 with respect to HepG2 cells, when evaluated on Vero cell its TC50 to IC50 ratio was below that threshold and thus its anti-parasitic activity could not be considered specific (Table 1).
In contrast to all the aforementioned results, hippeastrine 2 did show low toxicities against Vero [TC50 = 45.99 (6.32) µM] and HepG2 cells [TC50 = 128.1 (12.26) µM], and it indeed complied with the SI window > 10 against both cell lines (Table 1). It was the only compound that had a SI > 10 versus Vero cells (SI = 12.7; Table 1). In addition, hippeastrine 2 presented the highest SI against HepG2 cells (SI = 35.3), with a TC50 value similar to that previously reported by Weniger et al. (TC50 = 40 µg/mL; i.e. 126.85 µM) (43). We thus assessed in a subsequent anti-amastigote biological assay whether hippeastrine anti-T. cruzi activity was indeed specific against this replicative form of the parasite. It was found that the observed anti-amastigote activity (IC50) was 3.31 (0.39) µM, which was again within 5× that of the reference drug BNZ in the same assay [IC50 = 1.2 (0.22) µM] (Figure 5). Moreover, there would yet be a SI > 10 for hippeastrine 2 with respect to its anti-amastigote activity (SI = 13.9 against Vero cells, and 38.7 against HepG2 cells).
Cytotoxicity results together with the in vivo assays reported by Zhou et al. (38) may predict a low toxicity in future evaluation of hippeastrine 2 anti-T. cruzi activity in animal models. Notwithstanding, before arriving at in vivo studies, other in vitro studies could be pursued to better qualify this alkaloid. For example, assessing its effect on T. cruzi CYP51 target (44) since it has been invalidated in the clinic (45, 46), and identifying whether it can kill dormant parasite forms (47). Moreover, it would be of interest to determine key in vitro pharmacokinetic (PK) parameters such as its solubility, permeability and clearance.
{kind=link}