Expression and diagnostic value of KIF23 in GC patients
The KIF23 expression level in tumor tissues was significantly higher than that in normal tissues (P < 0.001; Figure 1A), and also higher in tumor tissues than in paired normal tissues (P < 0.001; Figure 1B). To evaluate the diagnostic performance of KIF23 in GC, we conducted ROC curve analyses. The ROC analysis of TCGA-STAD revealed significant diagnostic accuracy with AUC=0.958 (95% CI 0.937–0.978) (Fig.2A). Thus, KIF23 had potential to be a novel diagnostic biomarker for GC.
Verification of KIF23 expression and diagnostic value in GC
To validate the protein level of KIF23 in GC, we performed immunohistochemistry and found that the expression of KIF23 was elevated in GC tissues (Fig 3) compared with that in adjacent normal tissues.According to the KIF23 IHC staining, 20% (6/30) of adjacent GC tissues showed low expression of KIF23, while 3.33% (29/30) of GC tissues showed high expression of KIF23. The profile of KIF23 mRNA expression in GC and normal gastric tissues was analyzed based on GEPIA (P < 0.05; Fig 1D). Finally, GSE2685 from the GEO databases was analyzed to verify the expression of KIF23 in GC. The expression of KIF23 was higher in the tumor tissues compared to that in normal tissues (Fig 1C). ROC curves were constructed to evaluate the diagnostic value of KIF23 for GC. The area under the ROC curve of GSE2685 was 0.86793 (Fig 2B).
Associations of KIF23 expression level with clinicopathological variables
Clinicopathological characteristics of the GC patients are listed in Table 1. As Table 1 showed, KIF23 expression was remarkably positively associated with age (P=0.004), histological type (P=0.006), H pylori infection (P=0.030). No significant difference in KIF23 mRNA level was found in patients with pathological T stage (P=0.756), pathological N stage (P=0.904), pathological M stage gender (P=0.626), pathological stage (P=0.356), primary therapyoutcome (P=0.635), gender (P=0.776), residual tumor (P=0.777) and histologic grade (P=0.129).
KIF23 expression in the stratified GC population
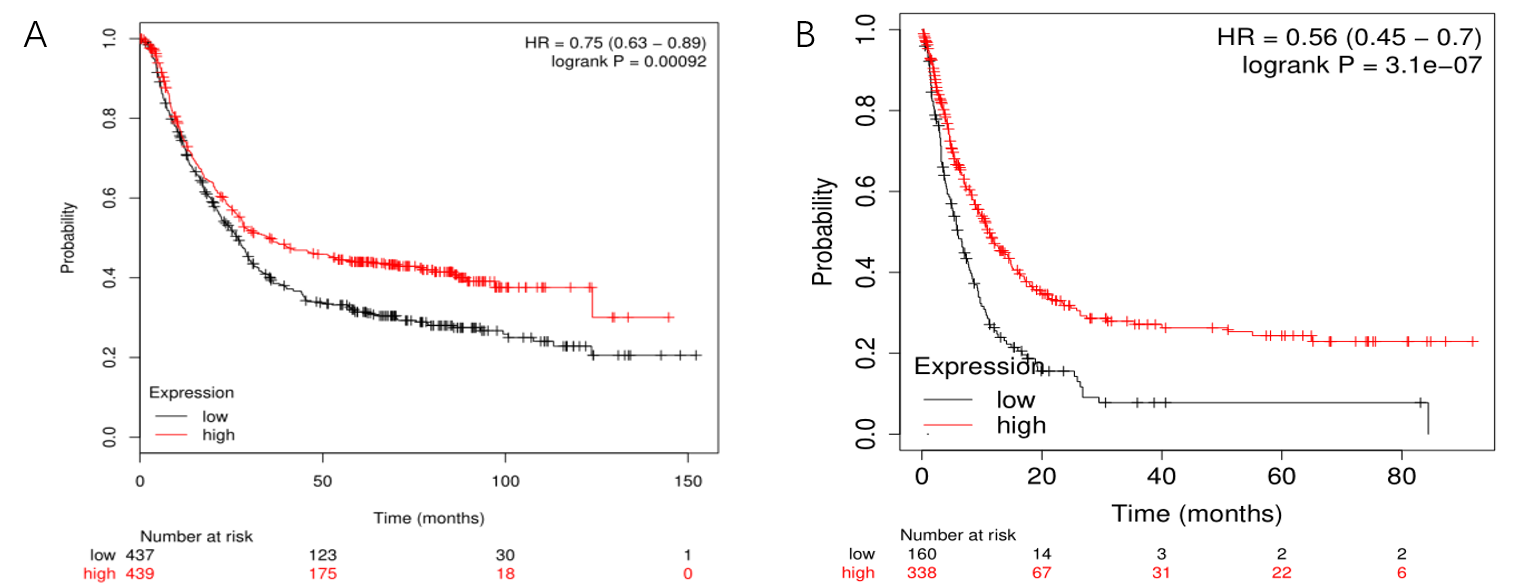
KIF23 expression was significantly correlated with OS and progress free interval (PFS) in GC patients (Fig.2C-D). Next, we analyzed the prognostic value of KIF23 through Kaplan–Meier plotter database. Notably, higher KIF23 expression was still correlated to better OS and PFS in GC (Supplementary Figure1). The results showed that higher KIF23 mRNA expression was significantly correlated with a longer prognosis.
Mutations of KIF23 in GC
The mutation frequency of KIF23 in GC was evaluated in the cBioPortal database. Five datasets (MSK, AMC, INSERM, RIKEN and TCGA-PanCancer Atlas), which included 1000 samples, were selected for analysis [18, 19]. The somatic mutation frequency of KIF23 in GC was 1.8%, which mainly consisted of missense mutations (Fig. 4A). This mutation frequency was relatively low, only 18 in 1000 samples. Furthermore, the mutation types of KIF23 were further evaluated in another database, COSMIC. For clarity, two pie charts of the mutation types are shown in Fig. 4. Missense substitutions occurred in approximately 42.39% of the samples, synonymous substitutions occurred in 11.11% of the samples, and frameshift deletions occurred in 11.36% of the samples (Fig. 4B). The substitution mutations mainly occurred at G>A (27.01%), followed by C > T (24.82%), C > A (10.22%) and G > T (9.85%) (Figure 4C).
Gene co-expression and hub gene analysis in GC
To further validate the biological activities of KIF23 in GC, the KIF23-related DEGs were evaluated in GC. The volcano map identified KIF23-related DEGs, with positively related genes on the right of the plot and negatively related genes on the left of the plot (Fig. 5C). Additionally, the heatmaps of the top 10 positively related genes were BUB1B, BUB1, PRC1, ARHGAP11A, C15orf23,TPX2, CCNB2, FANCI, NUSAP1 and ZWILCH (Fig. 5A). The top 10 negatively related genes identified were LTC4S, MARCH2, GYPC, FXYD1, CLEC3B, CBX7, JAM2, PBXIP1, GFRA1 and MFAP4 (Fig 5B). To determine the relationship of the top 100 positively related genes of KIF23 in GC, a PPI network was established. As shown in Fig. 4D, frequent interaction among the top 100 genes had close relationships with KIF23 expression. After calculating by degree using Cystoscope software, we obtained ten hub genes that revealed the closest relationships. The ten hub genes were BUB1, CDK1, CCNA2, CDCA8, CCNB1, CCNB2, KIF11, KIF2C, NCAPG and UBE2IR (Fig. 5E). Furthermore, Gene-MANIA tools were used to analyze the interaction between KIF23 and protein-coding genes mentioned earlier. Fig. 5F illustrates the results of the analysis.
Functional enrichment analysis and predicted signaling pathways
To better understand the functional implication of KIF23 in GC based on the top 100 significantly related genes, GO enrichment analysis was performed using the Cluster Profile package. GO results (Fig. 6A) revealed top four significant biological processes (BP), top four cellular components (CC), and top four molecular functions (MF). The results showed these co-expression genes were mainly involved in tubulin binding, microtubule and regulation of cell division in biological process, cellular component and molecular function, respectively. Moreover, according to KEGG analysis, the results of KIF23 related co-expression gene were mainly involved in the cell cycle, oocyte meiosis and secretion and DNA replication pathways (Fig. 6B). The results of KEGG pathway analysis showed that the functions of KIF23 and its neighboring genes were mainly enriched in the cell cycle, DNA replication, fanconi anemia pathway and homologous recombination (Fig. 6C). These results demonstrated that KIF23 has a wide range of effects on the genes and pathways involved in cell cycle.
Correlation between KIF23 expression and immune microenvironment in GC
In TIMER database, KIF23 expression was related with the purity in 16 cancer types, B cell infiltration in 24 cancer types, CD8+ T cell infiltration in 17 cancer types, CD4+ T cell infiltration in 17 cancer types, macrophage infiltration in 21 cancer types, neutrophil infiltration in 24 cancer types, and dendritic cell infiltration in 21 cancer types (Fig.7 and Supplementary Figure 2).
We next found that KIF23 was correlated with the infiltration of six types of immune cells (B cell, CD8+ T cells, CD4+ T cells, macrophage, neutrophil, DCs) in GC and ESCA (Fig. 7A). To be specific, KIF23 expression was negatively related to the infiltration of B cells (r=-0.162, P=1.79E-03), CD8+ T cells (r=-0.236, P=4.62E-06), CD4+T cells (r=-0.218, P=2.57E-05), macrophages (r=-0.324, P=1.65E-10), neutrophils (r=-0.132, P=1.09E-02), and dendritic cells (r=-0.233, P=5.63E-06) in GC. However, KIF23 expression was not significantly related to the infiltration of B cells (r=0.044, P=5.57E-01), CD8+ T cells (r=-0.046, P=5.39E-01), CD4+T cells (r=-0.139, P=6.42E-02) and macrophages (r=0.034, P=6.47E-01), except neutrophils (r=-0.191, P=1.02E-02) and dendritic cells (r=-0.196, P=8.27E-03) in ESCA. These findings suggested that KIF23 expression was correlated with immune infiltration in GC.
We further compared the infiltration level in GC and ESCA with different somatic copy number alterations (SCNA) in KIF23 (Fig.7B). In ESCA, CD8+ T cell was the main infiltrating immune cell in the arm-level deletion state of KIF23 gene, while B cell was the main infiltrating immune cell in the arm-level gain state (all P<0.05). In GC, B cell, CD8+ T cell, CD4+ T cell, macrophage, neutrophil and dendritic cell were the main infiltrating immune cells in the arm-level deletion state of KIF23 gene, while CD8+ T cell, CD4+ T cell, neutrophil and dendritic cell were the main infiltrating immune cells in the arm-level gain state (all P<0.05). B cell and CD8+ T cell were the main infiltrating immune cells in the high amplication state of KIF23 gene (all P<0.05).
Correlation analysis between KIF23 expression and immune marker sets
We analyzed the correlations between KIF23 and marker genes of different immune cells, including CD8+ T cells, T cells(general), B cells, monocytes, TAMs, M1 macrophages, M2 macrophages, neutrophils, NK cells and DCs in GC, using esophageal carcinoma (ESCA) as the control (Table 2 and Fig.8–9). Moreover, we analyzed the levels of functional T cells, namely, Th1 cells, Th2 cells, Tfh cells, Th17 cells, and (regulatory)Tregs, as well as exhausted T cells. After adjustment for purity, the results revealed the KIF23 expression was significantly correlated with the expression of markers of most immune cells and T cells in ESCA and GC. (Table 3). TIMER and GEPIA analyses showed that KIF23 expression in GC tissue was significantly correlated with the expression of marker genes in B cells, monocytes, TAMs, neutrophils, DCs, T-helper, Tfh, Treg and exhausted T cells, but not in ESCA (Fig 8-9and Table 3).
KIF23 expression showed significant correlation with the expression of markers of immune cells: B cell marker, CD19 (r=-0.168; P=1.03e-03), CD79A (r=-0.276;P=4.59e-08); monocyte marker, CD115 (r=-0.136; P=8.21e-03); TAM marker, CCL2 (r=-0.216;P=2.22e-05); M1 macrophage marker, iNOS (r=0.18; P=4.38e-04); M2 macrophage marker, MS4A4A (r=-0.127; P=1.35e-02); neutrophil markers, CD66b (r=0.175; P=6.16e-04) and CCR7 (r=-0.232; P=5.02e-06); NK cell markers, KIR2DL1(r=0.118; P=2.11e-02), KIR2DL3(r=0.121; P=1.81e-02) and KIR2DL4(r=0.213; P=2.95e-05); DC markers, BDCA-1 (r=-0.375,P=3.97E-14), BDCA-4(r=-0.3,P=2.69E-09).
Significant correlation existed between KIF23 and marker genes of exhausted Treg and T cells: Th1 marker, STAT1 (r=0.4, P=1.8E-17) and IFN-γ (r=0.19, P=8E-05); Th2 marker, GATA3(r=-0.16, P=1.4E-03); Tfh marker, BCL6(r=-0.1, P=0.038) and IL21(r=0.067, P=0.18); Th17 marker, IL17A (r=0.19, P=8.1E-05); Treg marker, TGFβ (r=-0.13, P=8.7E-03). Notably, exhausted T cell markers CTLA-4 (r=0.16, P=8.4E-04) and GZMB (r=0.16, P=1.5E-03) had a moderate correlation with KIF23 expression, suggesting that high KIF23 expression plays an important role in CTLA4-mediated T cell exhaustion. Therefore, these findings confirm that KIF23 is specifically correlated with immune infiltrating cells inGC.
{kind=link}