1. DHA is a promising candidate showing some therapeutic advantages in EAE
Firstly, we identified the therapeutic effect of DHA on the development and severity of mice with EAE. The onset of clinical signs began on the 4th day postimmunization. As time went on, in EAE group, all mice had developed clinical symptoms and the mean clinical score was 3.92 ± 0.25 on the 18th day at the peak of the disease, also accompanied by paralysis of all four limb. In contrast, the mean clinical score in the DHA 10 mg.kg− 1 treated group (1.98 ± 0.21) was significantly reduced to nearly two-fold. Strikingly, the disease score was even less than one-fold than treatment with MET (2.84 ± 0.19), the most widely used drug for MS in the clinical treatment (Fig. 1A). Besides, DHA treatment obviously prevented the body weight loss in EAE mice (Fig. 1B). These results predicted that DHA had therapeutic potential in preventing EAE.
Histologically, to make further effort to DHA efficacy verification, spinal cord and brain tissue sections were prepared and then H&E staining was performed. Results in Fig. 1C and D showed that DHA significantly inhibited the pathological infiltration of inflammatory cells. Then, to histologically detect the protective effect on myelin, we used Luxol Fast Blue (LFB) staining, a specific myelin detection method. In EAE group, result in Fig. 1E showed the intensity of LFB staining in corpus callosum was weakened, most myelin lipids were pale blue, and blue filaments were hardly visible at the edge of spinal cord which can be seen as marker of demyelination. By contrast, DHA treatment remarkably prevented the demyelination.
Morphologically, we directly observed the ultramicroscopic structure of pathological changes by using transmission electron microscopic. Result in Fig. 1F showed an "orbital like" compact lamellar structure with no shrinkage, tightly arranged myelin sheath in negative control group. The impaired axons and inner vacuoles were also rarely observed. On the contrary, in EAE group, there were obvious pathological changes in the myelin sheath, the shape of myelin sheath was irregular, the lamella of myelin sheath was obviously loose, disintegrated, fused, or even lost. Consistent with previous results, the damages in myelin, as quantified by demyelinating or intact areas of nerve fibers, could be significantly prevented after DHA 10 mg.kg− 1 treatment, which even showed more protective effects than MET group.
Of note, by using gait analysis, we for the first time verified the efficacy of DHA. Results were shown in Fig. 1G. DHA treatment can obviously reverse the disorder of walk speed, Lh-Rh Pressure, Lh-Rh support time, Lh-Rh Print area, and ameliorate Body Rotation Average. Furthermore, it showed more therapeutic effects than MET in the perspective of walk speed and Lh-Rh Pressure. In conclusion, results from different aspects convinced us the therapeutic significance of DHA on EAE mice, which supported its future application in MS.
2. AXL in microglia was the potential responsive molecule of DHA in EAE
To detect the drug-responsive cell subset in 10x Genomics, we firstly used tSNE (t-Distributed Stochastic Neighbor Embedding) projection. Then, we used Louvain optimization algorithm to cluster and classify the cell groups. The responsive cell-types to DHA can finally be categorized into 14 sub-populations. Among them, Macrophage and microglia were for the first time identified as the most responsive cells and illustrated by graph-based clustering (Fig. 2A).
Furthermore, we also geonomy-widely provided the description for the DHA responsive molecular network and screened the specific marker genes expression in microglia. Result indicated that AXL was specifically induced by DHA in microglia from EAE mice. Pathologically, gene deficiency of AXL in EAE would cause much more excessive autoimmune inflammatory reaction, consequently inducing myelin sheath damage [30]. This study gave us great inspiration and reliable evidence that AXL may be a potential target for DHA treatment (Fig. 2B).
3. The inflammatory–resolving effect of DHA is functionally correlated with AXL
To immunologically elucidate DHA mediated resolution of autoimmune inflammation, the AXL-concentrated efficacy analysis was functionally conducted mainly through three aspects. These included the regulation of phagocytic and chemotactic potential in microglia as well as the regulation in co-regulatory signaling during antigen-presentation.
Firstly, to identify the prediction from 10x Genomics, we quantified the transcription level of AXL in mononuclear cells (MNCS) collected from CNS tissues in EAE mice, whose total number reflects the degree of infiltrated inflammatory cells [21]. As shown in Fig. 3A, by qRT-PCR assay, DHA treatment can up-regulate the expression of AXL compared with the vehicle control group. In contrast, GAS6, the ligand for AXL, showed no differences among each group (Fig. 3B). Consistent with the transcriptional result, the protein expression of AXL was further detected. Results showed that (Fig. 3C and D), specifically under inflammatory condition, DHA can up-regulate AXL compared with modeling group.
As indicated by previous report, relying on AXL, microglia can sense and engulf apoptotic cells, clear the myelin debris in CNS. Facilitating by such mechanism, the over-activated inflammation can be rapidly resolved and effectively confined within a small area. We therefore, functionally evaluated the impact of DHA on phagocytosis of microglia in vitro. In this study, the 5-Fu induced apoptotic PC12 cells (the apoptosis rate: more than 81%, Fig. 3E) were co-cultured with BV2 cells and the phagocytotic intensities were quantified by flow cytometry and further visualized by microscopic observation. As shown in Fig. 3F and G, compared with LPS group, the count and percentage of PC12 cells phagocytized by BV2 cells in DHA (1.0/4.0 µM) groups were both significantly increased. This result proved that the phagocytic ability of microglia under inflammation condition can be significantly enhanced by DHA, forming the important basis for inflammation resolution.
In the next study, from the perspective of “antigen uptake and presentation”, we aim to reveal the influence of DHA on APCs. Inspired by the essential roles of co-regulatory factors during EAE progression, detection of PDL1 and CD80 were chosen as the markers for the immunological properties during antigen presentation. As shown in Fig. 4A and B, in both Raw264.7 cells and peritoneal macrophages, DHA obviously increased the transcription of PDL1 compared with LPS treated group. However, there are no changes in the transcription of CD80 between DHA and LPS treated groups. Consistently, Flow cytometry analysis showed that the enhanced surface expression of PDL1 can be clearly detected in DHA treated macrophages as well as BV2 microglia cell lines (Fig. 4C, D and E). These results provided molecular indications that, by upregulating PDL1 in microglia, DHA showed potential for enhancing the co-inhibitory signaling during antigen presentation and neutralizing the excessive inflammatory signals in the progression of autoimmune diseases.
To further detect whether DHA directly influences the differentiation of T cells, we established a co-culture model of BV2-Jurkat T cells. Results in Fig. 4F showed that DHA (1.0/4.0 µM) significantly elevated the percentage of Treg cells (CD4 + Foxp3+), compared with LPS group. These results showed that DHA suppressed autoimmune inflammation partially through enhancing of PDL1 expression and promoting the differentiation of Treg cells.
According to the indication from the 10x Genomics, CCL5 is one of the most responsive molecules in DHA treatment (Fig. 5A and B). Inspired by the crucial role of CCL5 in the trafficking of encephalitogenic T cells, we firstly analyzed the level of CCL5 by using the qRT-PCR and ELISA assays (Fig. 5C). In line with the geonomy-wide screening, DHA treatment can obviously inhibit the gene expression in MNCS separated from EAE mice. Besides, the high level of CCL5 in the cerebrospinal fluid (CSF) of EAE mice can also be significantly reversed by DHA treatment (Fig. 5D). which shows the same pattern in the serum (Fig. 5E).
To functionally determine the influence of DHA on the chemoattraction of microglia, we used LPS to induce the production of CCL5 for 12 hours. Next, the supernatant was collected for the detection of CCL5 concentration by using ELISA assay. The results in Fig. 5F showed that DHA treatment can obviously reduce LPS-stimulated CCL5 secretion. Trans-well assay showed that DHA (1.0/4.0 µM) dramatically reduced the chemotactic ability of BV2 cells in LPS treated model (Fig. 5G).
4. The inflammatory-resolving effects of DHA necessitated the AXL signaling in microglia
Molecularly, in order to clarify whether the inhibition of autoimmune inflammation of DHA is required for the presence of AXL. We used SGI7079, the phosphorylation blocker of AXL at Tyr702. Figure 6A demonstrated that challenging with SGI7079 successfully blocked phosphorylation of AXL and the related regulation on STAT1:SOCS3 pathway. In addition, Fig. 6 also implied SGI7079 is sufficient to neutralize the inflammation resolving effects of DHA in microglia. The T cell differentiation (Fig. 6B, C), phagocytic potential (Fig. 6D, E) and chemotactic sensitivity (Fig. 6F, G) consistently showed no statistical differences among DHA, LPS and NC treated group (P > 0.05), indicating that AXL would be a potential target of DHA in treating EAE.
5. DHA specifically rebalanced overactivated inflammation by regulating AXL related STAT1:SOCS3 pathway
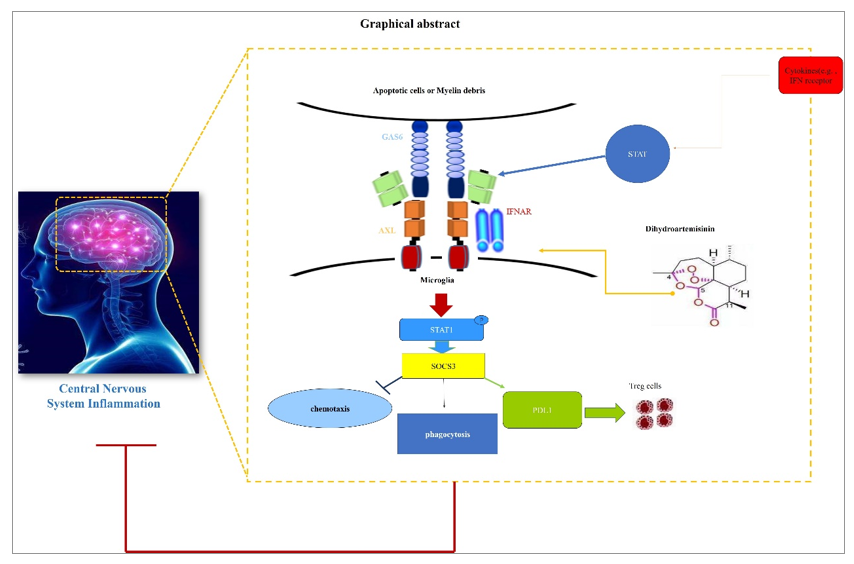
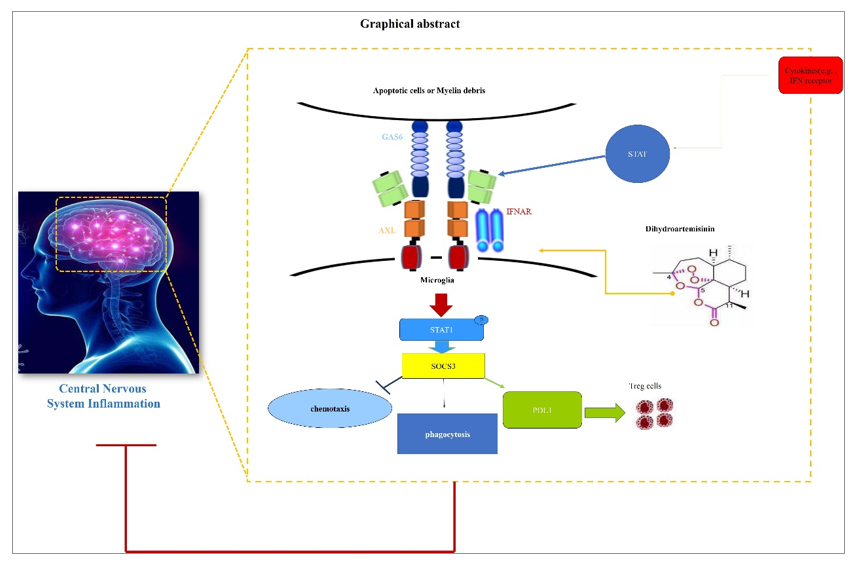
Large amounts of researchers have found that, specifically under the pro-inflammatory conditions, AXL interacted with type I interferon receptor (IFNAR) and potentiated the downstream inflammatory-resolving events as represented by the activation of STAT1:SOCS3 pathway. Of great interest, as the molecular switch for inflammation from promotion to resolution, the transcription of AXL itself is inversely facilitated by STAT1, a central inflammatory sensor regulated by various types of pro-inflammatory signals including IFN-β ligation to IFNAR. This forms the endogenous basis for the negative feedback loop of immune responses and consequently maintains the homeostasis in inflammatory microenvironment (graphically shown in below, Fig. 7.).
In light of this, to molecularly clarify the mechanistic revelation of DHA to IFNAR- STAT1 pathway, inflammatory stimulator to IFNAR (LPS or more specifically, IFN-β) were used and the influences of DHA on this pathway were then evaluated in BV2 cells. As shown in Fig. 8, in the presence of DHA, the expression of AXL can be dramatically induced by both LPS (Fig. 8A and D) and IFN-β (Fig. 8B and E), leading to a molecular pattern with higher inflammation-resolution potential, as suggested by SOCS3 upregulation. Moreover, in sharp contrast to the results obtained from the STAT1-avaliable state, the expression of AXL and SOCS3 were kept at the similar levels comparing to modeling group when phosphorylation of STAT1 was blocked by Fludarabine (FLU) (Fig. 8C and F). This result revealed that the enhanced AXL expression and the negative regulation of DHA in inflamed microglia were both required for the normal-responsiveness of STAT1 pathway.
Next, recent findings further proved that the inflammatory resolution effects of AXL shows high specificity to proinflammation conditions, which suggested that the biological activities of AXL were integrated into the IFN-β-IFNAR activation. To test such specificity, in the non-inflamed state, the STAT1:SOCS3 pathway was analyzed in the presence of DHA. In addition, the inflammatory resolving functions mediated by AXL were phenotypically measured in DHA treated BV2 cells. As shown in Fig. 9A, without inflammatory stimulation, DHA failed to regulate STAT1-SOCS3 pathway. Functionally, the differentiation induction of Treg cells (Fig. 9B, C), phagocytic (Fig. 9D) and chemotactic influences of BV2 cells (Fig. 9E), as above-proved manipulated by DHA, were totally neutralized. These results indicated that DHA functions as a resolution promoter by regulating AXL related STAT1:SOCS3 pathway specifically in the condition of over-activated inflammation.
{kind=link}
{kind=link}