Sequence correction of ERF and DREB subfamily genes
To ensure the sequence accuracy of all AP2/ERF genes, the Pfam model (pf00847) of the AP2 domain downloaded from the Pfam website was used to search the tomato v4.0 protein database. A total of 166 AP2/ERF proteins with an AP2 domain E-value < 0.001 were obtained. Among these proteins, 20 had ≥ 2 AP2 domains, while 146 proteins had single AP2 domain. Among the latter, 3 proteins with the B3 domain were RAV-type AP2/ERF proteins. Thus, there were 143 ERF subfamily proteins with a single AP2 domain. The 143 protein sequences were submitted to the Pfam, CDD, and smart websites for conservative domain analysis. Subsequently, 140 tomato ERF subunit genes with a single AP2 domain were identified. The sequences of these genes were compared in 3 tomato genome sequencing DNA, CDS, cDNA, and protein databases (versions 2.0, 3.2, and 4.0); 26 genes were found to be different (Table S1). The CDS and protein sequences of these 26 genes were compared and confirmed according to the tomato genome and NCBI databases (Tables S2 and S3). Finally, the corrected protein sequences were used for subsequent analyses (Tables S2).
Characteristics, polarity, and chemical structure analysis of the 14th and 19th amino acids in the AP2 domain
Among the 140 ERF genes with a single AP2 domain, the 14th amino acid of the AP2 domain was V in 57 genes. Among these 57 genes, the 19th amino acid of the AP2 domain was glutamic acid (E) in 30 genes, aspartic acid (D) in 4 genes, ssparagine (N) in 1 gene, glutamine (Q) in 4 genes, histidine (H) in 6 genes, leucine (L) in 10 genes, alanine (A) in 1 gene, and V in 1 gene (Tables 1 and S4). These 57 genes were identified as DREB genes. Additionally, the 14th and 19th amino acids of the AP2 domain were isoleucine (I) and D, respectively, in SlERF2-5, SlERF10-6, and SlERF10-8. The codon of I was AUA/AUC, GUA/GUG/GUU/GUC for V, but GCA/GCG/GCU/GCC for A. The characteristics, polarity, and chemical structure of I and V were hydrophobic, nonpolar, and aliphatic, while A was neutral, nonpolar, and aliphatic (Tables 1 and S4). Thus, I can only be a V mutation. Accordingly, the 3 genes were identified as DREB genes. In the 19th amino acid of the AP2 domain, the hydrophilic amino acids included E, D, N, Q, and H, the hydrophobic amino acids included L and V, and the neutral amino acids included A. The negative charged amino acids (E and D), uncharged amino acids (N and Q), and positively charged amino acids (H) were polar; the nonpolar amino acids included L, A, and V. Additionally, H had a heterocycle chemical structure, while the others were aliphatic (Table 1). These differences may affect the functionality of DREB protein interactions with DRE and GCC boxes.
Table 1
The 14th /19th amino acid analysis of the DREB subfamily AP2 domain
| Gene number | 14th | 19th | 14th codon | 19th codon | 14th /19th Characters | 14th polarity | 14th chemical structure | 19th polarity | 19th chemical structure |
| 30 | V | E | GUA/GUG/ GUU/GUC | GAA/ GAG | Hydrophobic/Hydrophilic | Nonpolarity | Aliphatic | Polarity with negative charge | Aliphatic |
| 4 | V | D | GUU | GAU/ GAC | Hydrophobic/Hydrophilic | Nonpolarity | Aliphatic | Polarity with negative charge | Aliphatic |
| 1 | V | N | GUU | AAC | Hydrophobic/Hydrophilic | Nonpolarity | Aliphatic | Polarity, without charge | Aliphatic |
| 4 | V | Q | GUA/GUG/ GUU | CAA | Hydrophobic/Hydrophilic | Nonpolarity | Aliphatic | Polarity without charge | Aliphatic |
| 6 | V | H | GUU/ GUC | CAU/ CAC | Hydrophobic/Hydrophilic | Nonpolarity | Aliphatic | Polarity with positive charge | Heterocycle |
| 10 | V | L | GUA/GUG/ GUU/GUC | UUG/CUU/ CUA/UUA | Hydrophobic/Hydrophobic | Nonpolarity | Aliphatic | Nonpolarity | Aliphatic |
| 1 | V | A | GUA | GCA | Hydrophobic/Neutral | Nonpolarity | Aliphatic | Nonpolarity | Aliphatic |
| 1 | V | V | GUG | GUU | Hydrophobic/Hydrophobic | Nonpolarity | Aliphatic | Nonpolarity | Aliphatic |
| 2 | I | D | AUA | GAC | Hydrophobic/Hydrophilic | Nonpolarity | Aliphatic | Polarity with negative charge | Aliphatic |
| 1 | I | V | AUC | GUU | Hydrophobic/Hydrophobic | Nonpolarity | Aliphatic | Nonpolarity | Aliphatic |
Among the 80 ERF subfamily members, the 14th and 19th amino acids of the AP2 domain were A and D in 70 genes. Additionally, there was an A and tyrosine (Y) in 1 gene, A and N in 1 gene, threonine (T) and D in 1 gene, serine (S) and D in 4 genes, E and D in 1 gene, glycine (G) and N in 2 genes, and I and V in 1 gene (Tables 2 and S5). In the 14th amino acid of the AP2 domain, the neutral amino acids included A, T, S, and G, and the hydrophilic amino acid included E. The nonpolar amino acid was A, the polar amino acids without charges were T, S, and G, and the chemical structure of these amino acids is aliphatic. In the 19th amino acid of the AP2 domain, D, Y, and N comprised the hydrophilic amino acids, the negative charged amino acids (D), the uncharged amino acids (Y and N) were polar, and the chemical structure of these amino acids was aliphatic (Table 2). Thus, the 80 genes with a single AP2 domain were identified as ERF subfamily members. These differences may affect the functionality of ERF protein interactions with GCC boxes.
Table 2
The 14th /19th amino acid analysis of the ERF subfamily AP2 domain
| Gene number | 14th | 19th | 14th codon | 19th codon | Characters | 14th polarity | 14th chemical structure | 19th polarity | 19th chemical structure |
| 70 | A | D | GCA/GCG/ GCU/GCC | GAU/GAC | Neutral/Hydrophilic | Nonpolarity | Aliphatic | Polarity with negative charge | Aliphatic |
| 1 | A | Y | GCA | UAU | Neutral/Hydrophilic | Nonpolarity | Aliphatic | Polarity without charge | Aromatic |
| 1 | A | N | GCU | AAU | Neutral/Hydrophilic | Nonpolarity | Aliphatic | Polarity without charge | Aliphatic |
| 1 | T | D | ACG | GAU | Neutral/Hydrophilic | Polarity without charge | Aliphatic | Polarity with negative charge | Aliphatic |
| 4 | S | D | UCU/ UCA | GAU/GAC | Neutral/Hydrophilic | Polarity without charge | Aliphatic | Polarity with negative charge | Aliphatic |
| 1 | E | D | GAA | GAU | Hydrophilic/Hydrophilic | Polarity with negative charge | Aliphatic | Polarity with negative charge | Aliphatic |
| 2 | G | N | GGA | AAC | Neutral/Hydrophilic | Polarity without charge | Aliphatic | Polarity without charge | Aliphatic |
Phylogenetic analysis of ERF and DREB proteins
To understand their genetic relationships, the protein sequences of the 60 DREB and 80 ERF subfamily members were classified into 6 groups (Fig. 1). The I group included 51 DREB proteins. Among these proteins, 37 and 14 proteins differentiated into the I-A and I-B subgroups, respectively. The I-A subgroup included 30 proteins with V14E19, 3 proteins with V14Q19 (SlERF12-9, SlERF1-13, and SlERF7-1), 1 protein with V14A19 (SlERF11-4), 1 protein with V14V19 (SlERF1-5), and 2 proteins with V14L19 (SlERF6-5 and SlERF12-3) (Table S4). Seven CBF proteins (SlERF3-7, SlERF3-22, SlERF3-6, SlERF8-2, SlERF8-3, SlERF12-11, and SlERF1-3) clustered together and were in the I-A subgroup (Fig. 1). Additionally, the I-A subgroup included 4 repressor proteins (SlERF9-1, SlERF2-10, SlERF4-10, and SlERF4-11) with EAR motif (DLNxxP or LxLxL) (Table S4). However, the I-B subgroup only included 6 proteins with V14H19 and 8 proteins with V14L19. SlERF9-10 and SlERF8-14 were repressor proteins in the I-B subgroup (Fig. 1; Table S4).
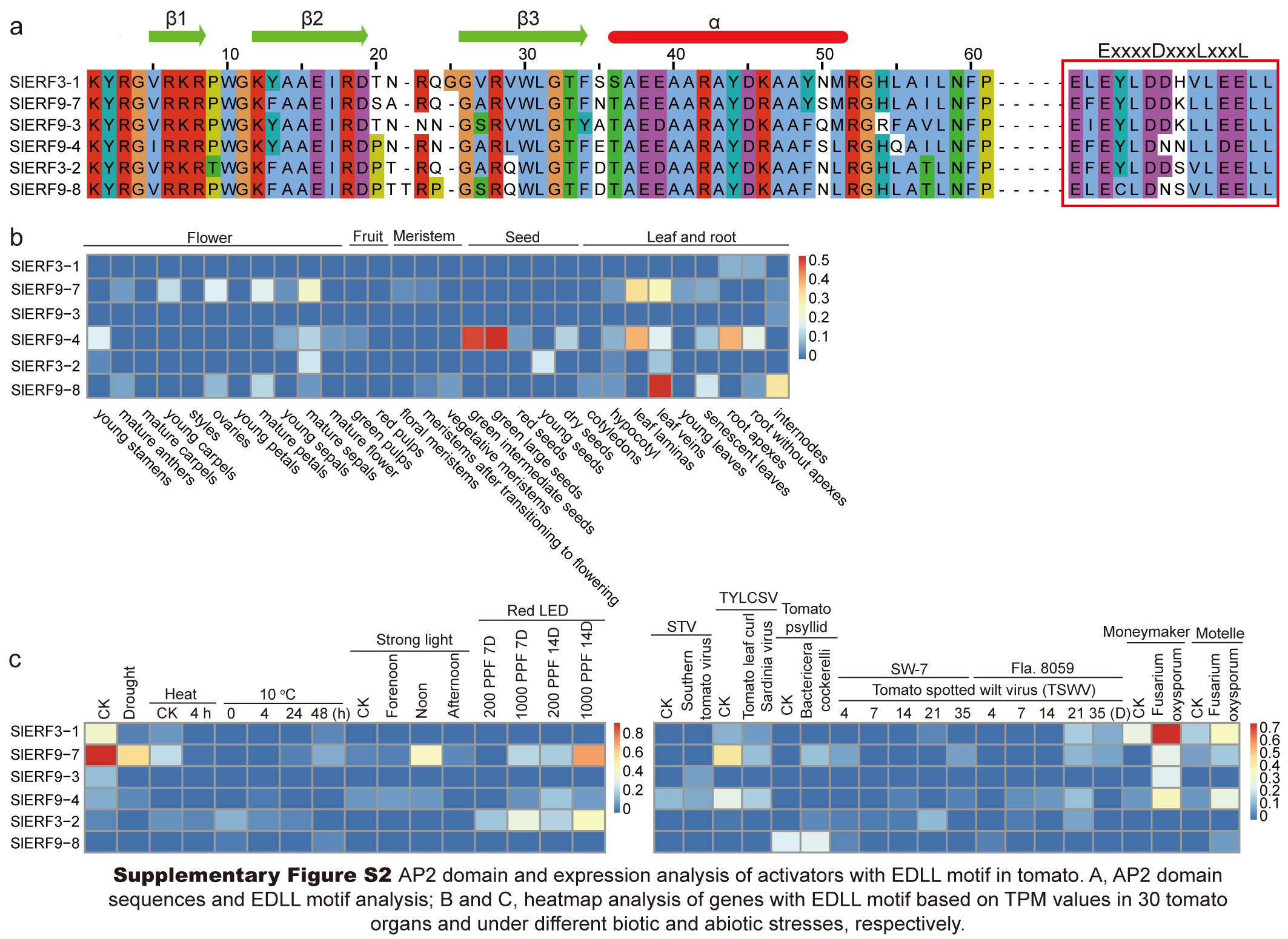
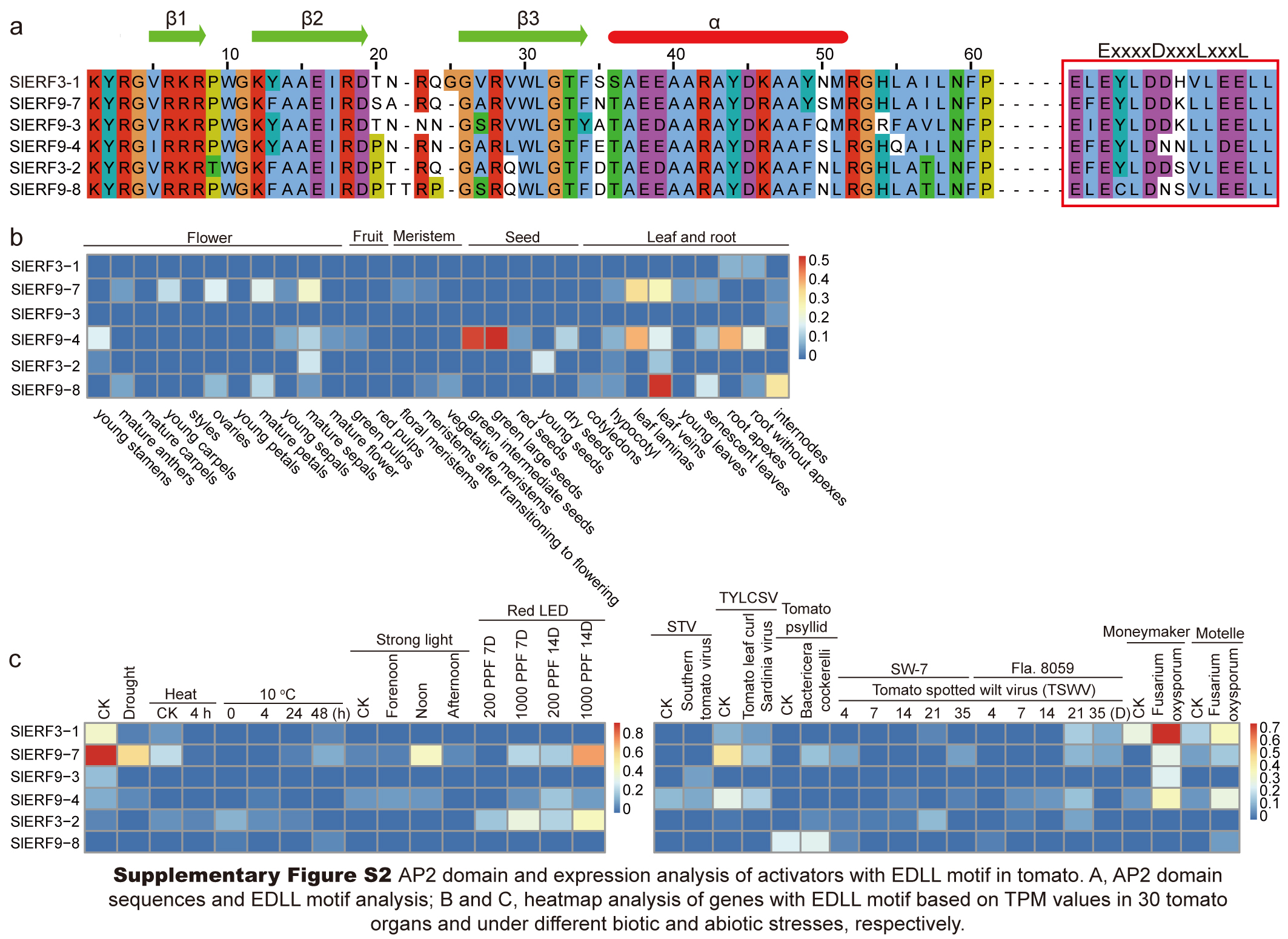
Group II included 49 ERF subfamily proteins. Among these proteins, 14 and 35 proteins differentiated into the II-A and II-B subgroups, respectively (Fig. 1). All members of the II-A subgroup belonged to ERF proteins with A14D19. In the II-B subgroup, there were 33 proteins with A14D19, 1 protein with T14D19 (SlERF1-10), and 1 protein with S14D19 (SlERF1-11) (Table S5). In the II-B subgroup, 6 proteins with the EDLL transactivation motif (ExxxxDxxxLxxxL) clustered together (SlERF3-1, SlERF9-7, SlERF9-3, SlERF9-4, SlERF3-2, and SlERF9-8) (Fig. 1). SlERF5-8 was also a repressor protein in the II-B subgroup. Groups III and IV included 13 and 16 ERF subfamily proteins, respectively. However, group III also included a DREB protein with V14D19 (SlERF1-4) that clustered with an ERF-type protein (SlERF1-2). One protein with S14D19 (SlERF1-15) and 1 repressor protein (SlERF4-1) were clustered into group III. Among these proteins in group IV, there were 2 proteins with S14D19 (SlERF3-16 and SlERF12-1), 2 proteins with G14N19 (SlERF12-6 and SlERF12-7), and all others belonged to proteins with A14D19. Additionally, 9 repressor proteins (SlERF10-1, SlERF7-5, SlERF12-1, SlERF7-2, SlERF2-6, SlERF7-3, SlERF10-2, SlERF3-4, and SlERF3-16) were in group IV (Fig. 1; Table S5). Group V had 8 DREB proteins, including 3 proteins with V14D19, 1 protein with V14Q19, 1 protein with V14N19, 2 proteins with I14D19, and 1 protein with I14V19 (Fig. 1; Table S4). Group VI included 1 protein with E14D19 (SlERF2-1) and 1 protein with A14D19 (SlERF9-2) (Table S5).
Motif analysis of ERF and DREB protein sequences
To understand the constructional characteristics of ERF and DREB proteins, a Multiple Em for Motif Elicitation (MEME) analysis was conducted to calculate the possible motifs of the 140 proteins. β1 of the AP2 domain was located on the left of motif 2, β2 was located on the right of motif 2 and left of motif 3, and β3 and α were located on motif 1. All 140 proteins, except SlERF9-1, SlERF10-9, SlERF6-1, SlERF8-4, SlERF8-12, SlERF3-8, SlERF2-1, and SlERF9-2, had motifs 1, 2, and 3. SlERF9-1, SlERF10-9, SlERF6-1, SlERF8-4, and SlERF8-12 had only motifs 1 and 3, as well as a same sequence to motif 25 in front of motif 3. SlERF3-8 had motifs 2, 3, and 16. Motif 16 had a similar sequence as motif 1. SlERF2-1 and SlERF9-2 did not have motif 1, 2 and 3, but had a similar sequence as motif 16 with motif 1 (Fig. 2). These results suggest that SlERF9-1, SlERF10-9, SlERF6-1, SlERF8-4, SlERF8-12, SlERF3-8, SlERF2-1, and SlERF9-2 may have the low ability to bind with GCC or DRE boxes.
In addition to motifs 1, 2, 3, 16, and 25, some motifs were located on both sides of the AP2 domain of many ERF and DREB proteins. For example, motifs 10 and 20 were near the left of the AP2 domain in 7 CBF and SlERF1-13, respectively, while motifs 4 and 5 were near the right of motif 1 and especially motif 4 (Fig. 2). These findings suggest that motifs 4, 5, 10, and 20 may be involved in the process of AP2 domain binding with GCC or DRE boxes. However, motifs 6–9, 11–15, 17–19, 21, 23, and 24 were relatively far away from the AP2 domain, may be located in the transactivation or repression domains, and may be involved in regulating the expression of their downstream target genes. However, some ERF and DREB proteins, including SlERF4-10, SlERF7-1, SlERF10-5, SlERF6-7, SlERF3-15, SlERF6-9, SlERF3-16, SlERF12-6, SlERF12-7, SlERF3-4, SlERF7-3, SlERF2-6, SlERF3-13, and SlERF2-5, did not have other motifs, except motifs 1, 2, 3, and 4 (Fig. 2). Nevertheless, a few of these proteins had a typical EAR (LxLxL or DLNxxP) repression domain, including SlERF4-10, SlERF3-16, SlERF3-4, SlERF7-3, and SlERF2-6. These proteins bound to DNA with the AP2 domain and repressed the expression of downstream target genes with the EAR domain. However, SlERF7-1, SlERF10-5, SlERF6-7, SlERF3-15, SlERF6-9, SlERF12-6, SlERF12-7, SlERF3-13, and SlERF2-5 especially protein sequences with < 100 amino acids (SlERF12-6 and SlERF12-7) may competitively inhibit other ERF and DREB proteins (Fig. 2).
Unique amino acids affected the ability of protein to bind with DRE and GCC boxes
Previous studies found that some DREB and ERF subfamily proteins only bound to DRE or GCC boxes, but most of these proteins can also interact with these boxes. However, the correlation between the characteristics and binding ability of DREB and/or ERF subfamily proteins remains unclear. To distinguish the difference between DREB and ERF proteins during binding with DRE or GCC boxes, the AP2 domain amino acid sequences of 49 Arabidopsis and 19 other species ERF proteins, including 8 tomato ERF proteins, were compared. The binding assays of the 68 ERF proteins with DRE and GCC boxes were completed through an electrophoretic mobility shift assay (EMSA), yeast one-hybrid, or proteome chip assays. Among these proteins, there were 42 protein AP2 domains that included P9, 5 included H9, 5 included S9, 6 included N9, 3 included Q9, 2 included K9, 2 included T9, and 1 included I9 (Fig. 3). Only 19 proteins bound with GCC box, including 17 ERF with P9, 2 DREB with 1 P9 and 1 H9. Additionally, 37 proteins bound with DRE and GCC boxes, including 23 ERF with P9, 14 DREB with 1 P9, 4 H9, 4 N9, 2 Q9, 1 T9, 1 K9, and 1 I9. Only 12 proteins bound with DRE, including 1 ERF with P9, 11 DREB with 5 S9, 2 N9, 2 K9, 1 Q9, and 1 T9 (Fig. 3). These results suggest that almost all ERFs with P9 and H9 can interact with GCC box, and most can also bind with DRE. All DREB with S9 can only interact with DRE, but other DREBs with N9, K9, Q9, T9, and I9 may only bind with DRE or with DRE and GCC boxes. The A14 and A15 amino acids of ERF AP2 domain were conserved, but the 13th amino acid may be Y, F, or W. The W13 and V14 amino acids of the DREB AP2 domain were conserved, but the 15th amino acid may be S, A, or C (Fig. 3). These characteristics of ERFs and DREBs may affect the ability of proteins to bind with DRE and GCC boxes.
In tomato DREB subfamily members, there are 10 DREBs with S9W13V14S15 (SlERF3-20, SlERF5-5, SlERF6-8, SlERF8-10, SlERF8-11, SlERF8-12, SlERF9-1, SlERF10-9, SlERF11-4, and SlERF12-9) and 1 DREB with S9W13V14C15 (SlERF8-4) (Table S1), which suggests that these 11 DREBs may only bind with DRE. There was 1 DREB with S9W13I14A15 (SlERF10-6). However, the ability of this protein to bind with DRE and GCC boxes has not been determined. There were 4 DREBs with H9W13V14S15 (SlERF12-4, SlERF6-7, SlERF3-15, and SlERF6-9) and 9 DREB with H9W13V14A15 (SlERF3-14, SlERF9-10, SlERF8-5, SlERF8-14, SlERF4-6, SlERF12-13, SlERF4-9, SlERF7-4, and SlERF12-5) (Table S1). DREB with H9W13V14A15 can interact with GCC or both DRE and GCC boxes. Seven CBF proteins exist in tomato DREB subfamily members, including 5 CBF with N9W13V14C15 (SlERF3-7, SlERF3-22, SlERF3-6, SlERF8-2, and SlERF1-3) and 2 CBF with D9W13V14C15 (SlERF8-3 and SlERF12-11). There were 3 DREB with N9W13V14S15 (SlERF6-1, SlERF8-6, and SlERF1-13), 5 DREB with K9W13V14S15 (SlERF11-5, SlERF11-6, SlERF1-5, SlERF6-5, and SlERF12-3), 4 DREB with K9W13V14A15 (SlERF2-10, SlERF4-10, SlERF7-1, and SlERF1-4), 6 DREB with T9W13V14A15 (SlERF10-4, SlERF10-5, SlERF10-7, SlERF4-4, SlERF4-11, and SlERF5-11), 3 DREB with 1 Q9W13V14S15 (SlERF6-4), 1 I9W13V14A15 (SlERF6-2), and 1 A9W13V14S15 (SlERF12-2), 5 DREB with P9W13V14A15 (SlERF2-2, SlERF1-1, SlERF3-13, SlERF1-14, and SlERF3-19), and 1 DREB with P9W13I14A15 (SlERF10-8) (Table S1). These DREB proteins may interact with DRE, some of which may also bind with GCC boxes.
In tomato ERF subfamily members, there were ERFs with 22 P9Y13A14A15, 21 P9F13A14A15, and 21 P9W13A14A15 (Table S1). There were ERFs with K9Y13A14A15 (SlERF3-18), P9Y13A14S15 (SlERF5-7), P9Y13G14A15 (SlERF12-7), P9Y13G14V15 (SlERF12-6), T9F13A14A15 (SlERF3-2), K9F13A14T15 (SlERF5-1), Q9F13S14A15 (SlERF1-11), P9F13S14A15 (SlERF12-1), Q9F13T14A15 (SlERF1-10), E9W13A14A15 (SlERF1-2), 2 with K9W13A14A15 (SlERF4-1 and SlERF5-2), and 2 with P9W13S14A15 (SlERF1-15 and SlERF3-16) (Table S1). This indicates that P9A14A15 of the ERF AP2 domain may play an important role in binding with GCC box. However, it has not been determined why most ERF proteins can also interact with DRE in comparison with these ERF proteins that only bind with GCC boxes.
Expression analysis of CBF genes in tomato
To understand the function of different ERF and DREB genes in tomato growth and development, the expression levels of ERF and DREB subfamily members were analyzed according to tomato RNA-Seq data abtained from NCBI SRA library. Seven tomato CBF proteins had the PKKPAGR motif in the N-terminal and the DSAWR motif in the C-terminal of the AP2 domain, but these highly similar proteins to CBF did not have the same motif (Fig. 4a). CBF proteins had N/D9W13V14C15 in β2, while other proteins had N/S/A9W13V14S15 (Fig. 4a). This analysis suggests that CBF and its highly similar proteins may process different abilities with GCC and/or DRE boxes, which leads them to play different roles in regulating tomato growth, development, and defense responses to abiotic stress.
In different tomato organs, SlERF8-2 was mainly expressed in mature petals and sepals, cotyledons, hypocotyl, and leaf lamina and veins; SlERF8-3 was only expressed in mature petals and sepals, and hardly in other organs. SlERF12-11 and SlERF1-3 were hardly expressed in all organs, except SlERF1-3 in young stamen. SlERF3-6 was mainly expressed in mature sepals, hypocotyl, leaf lamina and veins, and internodes. SlERF3-7 was expressed in mature anthers and petals, young and mature sepals, meristems after transitioning to flowering, cotyledons, hypocotyl, leaf lamina and veins, root apexes and roots without apexes, and internodes, especially in cotyledons, hypocotyl, and leaf lamina. SlERF3-22 was expressed in mature anthers, petals and sepals, meristems after transitioning to flowering, cotyledons, hypocotyl, leaf lamina, and internodes (Fig. 4b; Data S1). These results indicate that 6 CBF genes, except SlERF12-11, may be involved in regulating the development of some floral organs, meristems after transitioning to flowering, cotyledons, hypocotyl, leaves, roots, and internodes. Although other highly similar DREB proteins to CBF do not have PKKPAGR and DSAWR motifs, most of these proteins play roles in some tomato organs. For example, SlERF6-1 was expressed in mature petals and sepals, meristems after transitioning to flowering, cotyledons, hypocotyl, and leaf veins. SlERF8-12 and SlERF8-11 were highly expressed in mature anthers and young sepals, respectively, but were lowly expressed or undetectable in other organs. SlERF3-20 was highly expressed in some organs, including mature anthers, floral meristems, meristems after transitioning to flowering, vegetative meristems, hypocotyls, leaf veins, young leaves, and internodes; SlERF6-8 was highly expressed in hypocotyls and internodes. These results indicate that CBFs and some DREB genes that are highly similar to CBF regulate the development of the same organs, such as the expression of SlERF8-2, SlERF3-7, SlERF3-22, SlERF6-1, and SlERF3-20 in meristems after transitioning to flowering (Fig. 4b; Data S1).
Among 7 CBF genes, SlERF12-11 and SlERF1-3 did not respond to abiotic stress, including drought, heat, cold, strong light, and red light. SlERF8-2 and SlERF8-3 positively responded to cold and red light, but not to drought, heat, or strong light. SlERF3-6 was positively induced by heat and red light, negatively regulated by drought, and did not respond to cold or strong light. SlERF3-7 was positively induced by heat, cold, and strong and red lights, but did not change due to drought. SlERF3-22 responded to cold, and strong and red lights, but did not change due to drought or heat (Fig. 4c; Data S2). These results suggest that different CBF genes may play different roles in response to different abiotic stressors. However, some DREB genes that are highly similar to CBF also responded to abiotic stress, including SlERF6-1, SlERF1-13, SlERF12-9, and SlERF6-8, which up-regulated by drought, and SlERF6-1, SlERF8-4, SlERF8-12, and SlERF12-2, which up-regulated by cold. SlERF9-1 and SlERF10-9 were not expressed or induced by abiotic stress, while SlERF8-10 and SlERF8-11 were lowly expressed under abiotic stress (Fig. 4c; Data S2).
Under biotic stress, among 7 CBF genes, SlERF8-2, SlERF3-6, SlERF3-7, and SlERF3-22 were up-regulated by the tomato spotted wilt virus (TSWV), did not respond to the southern tomato virus (STV), tomato leaf curl Sardinia virus (TYLCSV), and tomato psyllid (Bactericera cockerelli), and were down-regulated by Fusarium oxysporum. SlERF8-2 was down-regulated by F. oxysporum and did not respond to other biotic stressors. SlERF12-11 and SlERF1-3 were not expressed by any biotic stress (Fig. 4c; Data S3). Among the DREB genes that were highly similar to CBF, SlERF1-13, SlERF5-5, SlERF12-2, and SlERF6-8 only responded to TSWV. SlERF6-1 and SlERF8-12 were only down-regulated by F. oxysporum and hardly responded to biotic stress. Other DREB genes were hardly regulated by any biotic stressor (Fig. 4c; Data S3).
Expression analysis of repressor genes in tomato
Among the 140 ERF proteins, 11 ERF and 6 DREB subfamily proteins, which have 1 or 2 typical EAR (LxLxL or DLNxxP) motifs, were found. SlERF10-1, SlERF7-5, SlERF12-1, SlERF7-2, SlERF4-11, SlERF9-1, SlERF2-10, and SlERF4-10 had a DLNxxP motif in the C-terminal, SlERF2-6, SlERF3-16, SlERF8-14, and SlERF4-1 had an LxLxL motif in the C-terminal, and SlERF9-10 and SlERF5-8 had an LxLxL motif in the N-terminal. SlERF7-3, SlERF10-2, and SlERF3-4 had an independent LxLxL motif and DLNxxP motif in the C-terminal. The DLNxxP motif of SlERF7-3 and SlERF10-2 connected with LxLxL sequences and formed a strong repressive motif (Fig. 5a).
To understand the function of the 17 repressor genes, the expression levels of these genes were analyzed in tomato flowers, fruits, meristems, seeds, leaves, and roots. Among these genes, SlERF10-1, SlERF7-5, SlERF12-1, SlERF7-2, SlERF7-3, SlERF10-2, and SlERF2-10 were largely expressed in almost all organs. SlERF2-6, SlERF3-4, SlERF4-10, and SlERF5-8 were expressed in most organs, including flowers, seeds, and leaves. SlERF2-6 and SlERF3-4 were not expressed in the fruits, in contrast to SlERF4-10 and SlERF5-8. SlERF3-16 was only expressed in the floral and vegetative meristems, seeds, and young leaves. SlERF4-11 was only expressed in the seeds. SlERF8-14 and SlERF4-1 were lowly expressed in almost all organs. SlERF9-10 and SlERF9-1 were not expressed in almost all organs (Fig. 5b; Data S1). These results indicated that SlERF10-1, SlERF7-5, SlERF12-1, SlERF7-2, SlERF7-3, SlERF10-2, and SlERF2-10 were involved in regulating the development of almost all organs, while other repressor genes, except SlERF9-10 and SlERF9-1, played roles in regulating the development of some organs.
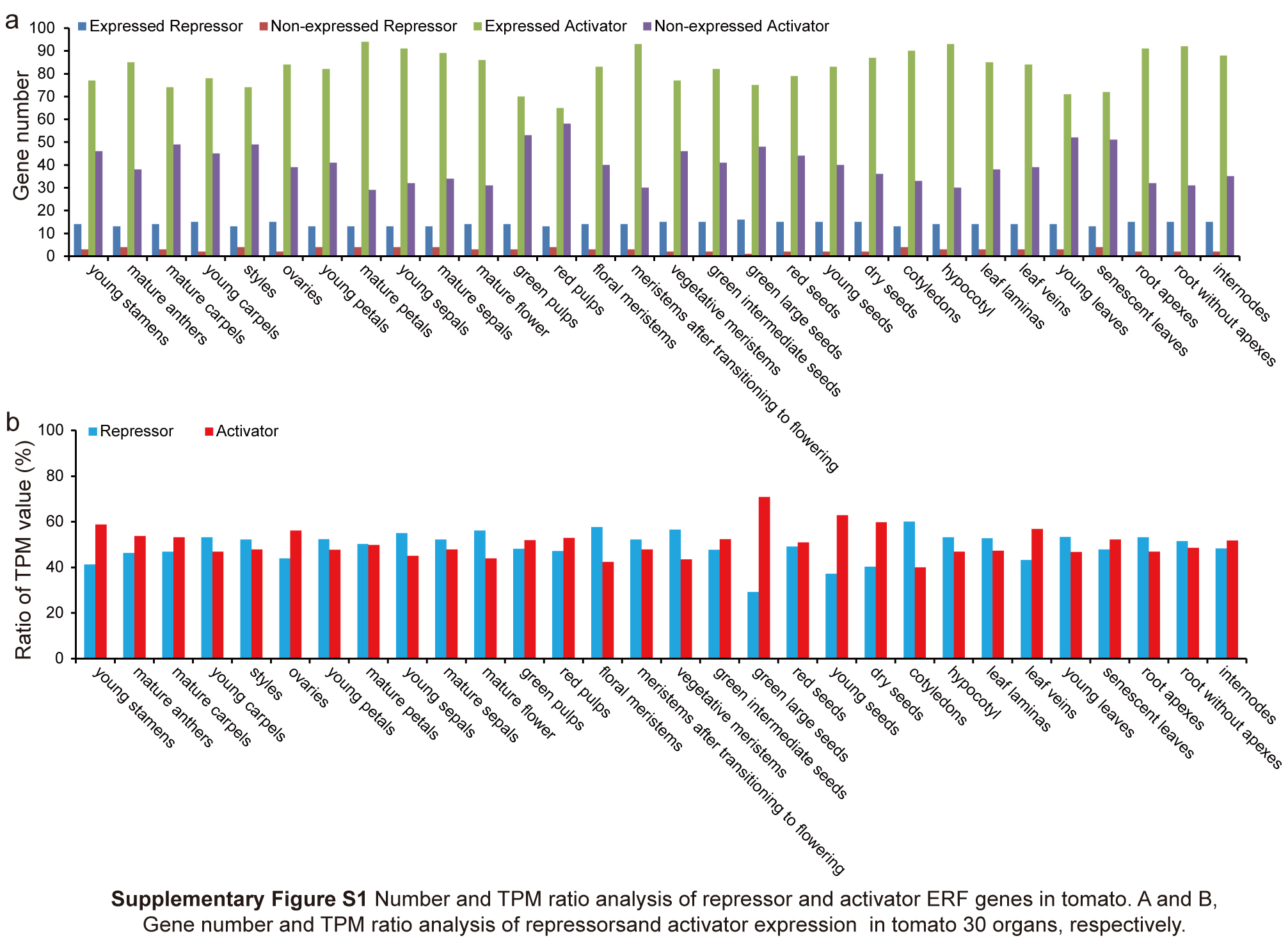
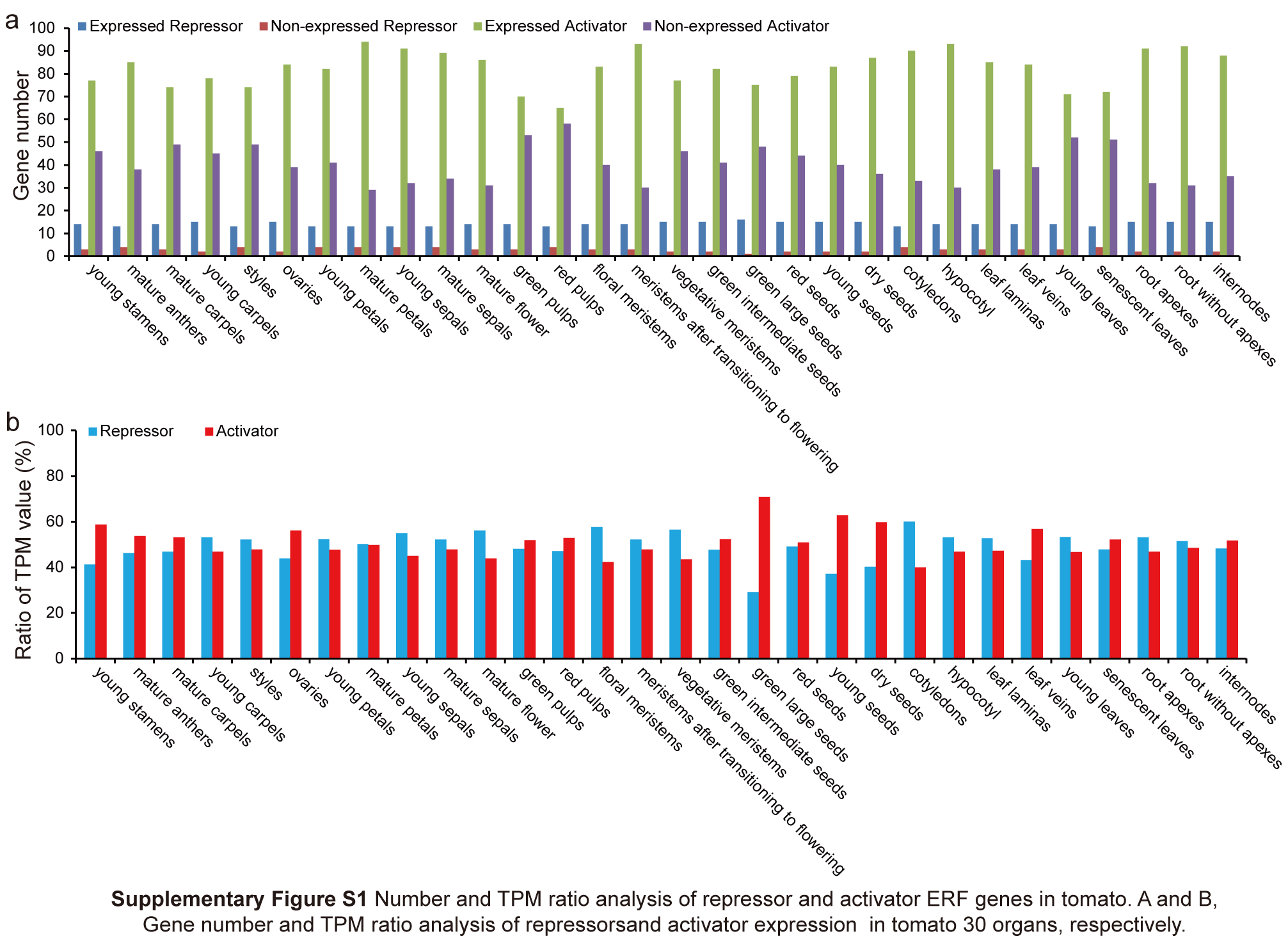
Among the 17 repressor genes, at least 13 were expressed in every organ, especially 16 in green large seeds (Fig. S1a; Data S1). Most activator genes were expressed in every organ, including a maximum of 94 genes in mature petals and minimum of 65 in red pulp (Fig. S1a; Data S1). However, the total TPM values of 17 repressor genes had a very high ratio among all ERF genes in every organ and exceeded the activator genes in most organs, including 56.14% in mature flowers, 57.64% in floral meristems, 56.58% in vegetative meristems, and 60.05% in cotyledons (Fig. S1b; Data S1). These results suggested that the 17 repressor genes play important roles in balancing the regulatory functions of other ERF and DREB subfamily genes and their downstream target genes during tomato growth and development.
Under drought stress, the expression of SlERF3-4 and SlERF4-10 increased, but other repressor genes did not change compared to the control (CK). The expression of SlERF10-1, SlERF2-6, SlERF10-2, SlERF5-8, and SlERF4-1 increased under heat stress, in contrast to SlERF8-14 (Fig. 5c; Data S2). SlERF7-3, SlERF10-2, SlERF8-14, SlERF5-8, and SlERF4-1 responded to cold stress, but other genes were not induced. Only SlERF10-2 was positively regulated by strong light at forenoon and SlERF8-14 was negatively regulated under strong light. SlERF10-1, SlERF2-6, SlERF7-3, SlERF10-2, SlERF2-10, and SlERF4-10 responded to red light stress (Fig. 5c; Data S2). A handful of ERF repressor genes responded to some biotic stressors, including SlERF10-1, SlERF7-5, SlERF7-3, and SlERF10-2, which were up-regulated after induction with TSWV in tomato species SW-7 and Fla.8059. SlERF2-6 was down-regulated by the STV, in contrast to the TYLCSV. SlERF8-14 was down-regulated by the TSWV and F. oxysporum. However, most ERF repressor genes, including SlERF7-2, SlERF3-16, SlERF9-10, SlERF4-11, SlERF9-1, SlERF2-10, and SlERF4-10, did not respond to the STV, TYLCSV, tomato psyllid, TSWV, or F. oxysporum (Fig. 5c; Data S3). These results suggest that some repressor genes are involved in regulating tomato tolerance to abiotic and biotic stress.
Expression analysis of ERF genes in the II-A subgroup
Among the 14 ERF subfamily genes in the II-A subgroup, the 5th, 23th, 24th, 46th, and 48th amino acids of the AP2 domain exhibited unique characteristics. SlERF4-2, SlERF4-5, SlERF4-7, SlERF12-12, SlERF3-17, SlERF6-10, SlERF12-8, SlERF1-16, and SlERF10-3 had V5A23A24L/I46F48, while SlERF1-6, SlERF9-6, SlERF12-10, SlERF3-21, and SlERF6-6 had I5G23V/I24R/K46I48 (Fig. 6a). These differences may affect the ability of ERF proteins to bind with DRE and GCC boxes during the regulation of growth, development, and tolerance to abiotic and biotic stress.
In 30 tomato organs, SlERF1-6, SlERF9-6, SlERF12-10, SlERF3-21, and SlERF6-6 were largely expressed in almost all organs, but SlERF4-7, SlERF12-12, SlERF3-17, SlERF12-8, SlERF1-16, and SlERF10-3 were only expressed in some organs, including SlERF4-7 in mature anthers, styles, and seeds, and SlERF12-12 in mature anthers, mature carpels, styles, ovaries, mature petals, mature sepals, and seeds. SlERF4-2, SlERF4-5, and SlERF6-10 exhibited low transcription levels in some organs and were not expressed in most organs (Fig. 6b; Data S1). These results suggest that most of the II-A subgroup genes play roles in regulating tomat growth and development.
All II-A subgroup genes did not respond to drought or strong light stress, but some of these genes were up-regulated by heat, cold, and red light. For example, the expression of SlERF4-2, SlERF1-16, and SlERF6-6 increased under cold and red light stress, and SlERF9-6, SlERF12-10, and SlERF6-6 increased under heat stress (Fig. 6c; Data S2). These results indicate that some of the II-A subgroup genes improve heat, cold, and red light tolerance. However, all II-A subgroup genes did not respond to biotic stress and some genes were even down-regulated, including SlERF1-16 and SlERF10-3 under F. oxysporum treatment (Fig. 6c; Data S3). These results suggest that the II-A subgroup genes were not involved in enhancing the tolerance to biotic stress.
Expression analysis of DREB genes in the I-B subgroup
Among the 14 DREB genes in the I-B subgroup, the 12th, 19th, 39th, 43th, 47th, 48th, and 49th amino acids of the AP2 domain exhibited unique characteristics. SlERF3-14, SlERF12-4, SlERF6-4, SlERF6-7, SlERF3-15, and SlERF6-9 had S12H19R39E43L47M48S/C49, while SlERF9-10, SlERF8-5, SlERF8-14, SlERF4-6, SlERF12-13, SlERF4-9, SlERF7-4, and SlERF12-5 had K12L19F/Y/L/M39T/R/K/V43M/K/R47L48R49 (Fig. 7a). SlERF9-10 and SlERF8-14 also show an EAR motif. These characteristics indicate that the ability of the I-B subgroup genes to bind with DRE and/or GCC boxes may be different, and these genes play different roles in regulating tomato growth and development.
Among the 14 DREB genes, SlERF4-6 and SlERF4-9 were highly expressed in all tomato organs, but other genes were specifically expressed in some organs, including SlERF3-14 in young stamens, young sepals, and senescent leaves, SlERF6-4 in young sepals and seeds, and SlERF3-15 in red and dry seeds. SlERF12-5 was not expressed in any tomato organs (Fig. 7b; Data S1). These results suggest that the I-B subgroup genes, except SlERF12-5, are involved in regulating different tomato growth and development processes.
Under abiotic stress, the expression of all I-B subgroup genes did not increase and SlERF8-5 was down-regulated by drought stress. Under heat stress, the expression of SlERF6-4 and SlERF8-14 decreased, SlERF4-9 and SlERF7-4 increased, and other genes did not change. SlERF8-14, SlERF4-9, and SlERF7-4 were up-regulated by cold stress, while other genes did not change. Almost all I-B subgroup genes did not respond to strong or red light stress, but SlERF3-14 and SlERF6-7 were positively regulated and SlERF6-4 was negatively regulated by red light (Fig. 7c; Data S2). All I-B subgroup genes did not positively respond to biotic stress (Fig. 7c; Data S3). These results indicate that a few I-B subgroup genes play roles in abiotic stress responses and all I-B subgroup genes are not involved in biotic stress responses.
{kind=link}
{kind=link}
{kind=link}
{kind=link}