Activation of the expression of the upstream regulator ACD6 of the SA pathway by drought stress
Our previous studies have shown the molecular mechanism underlying the induction of defense gene expression in the SA pathway by biotic stresses [4, 30]. To investigate whether abiotic stress could induce the expression of defense gene ACD6 in the SA pathway and stress resistance genes GSTF14 and ACO3, the wild-type Columbia (Col-0) line of Arabidopsis thaliana was selected for drought-stress treatment, cold-stress treatment, and salt-stress treatment. There were no significant phenotypic changes in plants treated with cold stress (4 °C) for 24 h or salt stress (150 mM) for 3 days. On days 5-7, the leaves of Col-0 plants treated with drought stress turned slightly yellow and shrunk (Figure 1B, C) in comparison to untreated Col-0 plants (Figure 1A). On day 14, anthocyanin accumulation in the leaves of Col-0 plants treated with drought stress clearly increased, and the leaves turned severely yellow and withered (Figure 1 D).
We extracted the total RNA from Col-0 plants on the 7th day of drought-stress treatment for comparative analysis of gene expression. The results of the reverse transcription–semiquantitative polymerase chain reaction (RT-sqPCR) assay showed higher expression levels of the regulator ACD6 of the SA pathway, the stress resistance gene GSTF14, and ACO3 in the plants after drought-stress treatment than in untreated Col-0 plants; GAPDH acted as a reference gene in this study (Figure 1E). Consistent with the RT-sqPCR results, the quantitative reverse transcription-polymerase chain reaction (RT-qPCR) analysis confirmed that ACD6, GSTF14, and ACO3 were significantly upregulated after drought-stress treatment, and the upregulation of GSTF14 expression was more significant (Figure 1F). Since ACD6 is an upstream regulator of the SA pathway, the increase in ACD6 expression could upregulate the expression of the defense genes NPR1 and PR5 (Figure 1G).
Figure 1 Detection and analyses of the expression of defense genes in Arabidopsis plants treated with drought stress
(A) The untreated Arabidopsis Col-0 plants. (B, C) The leaves of Arabidopsis plants treated with drought stress turned slightly yellow and shrunk during days 5-7. (D) Anthocyanin accumulation in the leaves of Arabidopsis plants treated with drought stress clearly increased, and the leaves turned severely yellow and withered on day 14. (E) Transcript levels of related genes in Arabidopsis plants treated with drought stress were analyzed by sqPCR; untreated Col-0 plants served as controls. (F) Transcript levels of related genes in Arabidopsis plants treated with drought stress were analyzed by qPCR. The statistical analysis was performed; asterisks indicate statistically significant differences compared with control plants (P < 0.05). (G) Transcript levels of defense genes in Arabidopsis plants treated with drought stress were analyzed by sqPCR.
Induction of SA pathway-related defense genes by cold and salt stress
To further investigate whether cold stress could also induce the expression of defense genes in the SA pathway, we extracted total RNA from wild-type Arabidopsis Col-0 plants treated under different conditions and detected the related defense genes. RT-sqPCR results showed that compared with controls, A. thaliana plants treated with cold or salt stress had significantly higher expression levels of defense genes ACD6, NPR1, and PR5 and ABA pathway-related gene ACO3 (Figure 2A, B). Consistent with the RT-sqPCR results, the RT-qPCR results further confirmed that cold stress and salt stress activated the expression of ACD6, which was significantly increased after 24 h of cold-stress treatment (Figure 2C, D). We also compared the expression of the stress resistance gene GSTF14. The results showed that the upregulation of GSTF14 was the most significant in the plants treated with cold stress for 24 h (Figure 2C).
Figure 2 Detection and analyses of the expression of defense genes and stress resistance genes in Arabidopsis
(A, B) The defense genes and ACO3 transcript levels in Arabidopsis plants treated with cold and salt stress were analyzed by sqPCR; untreated Col-0 plants served as controls. (C, D) ACD6, GSTF14, and ACO3 transcript levels in Arabidopsis plants treated with cold and salt stress were analyzed by qPCR; untreated Col-0 plants served as controls. The statistical analysis was performed; asterisks indicate statistically significant differences compared with control plants (P < 0.05).
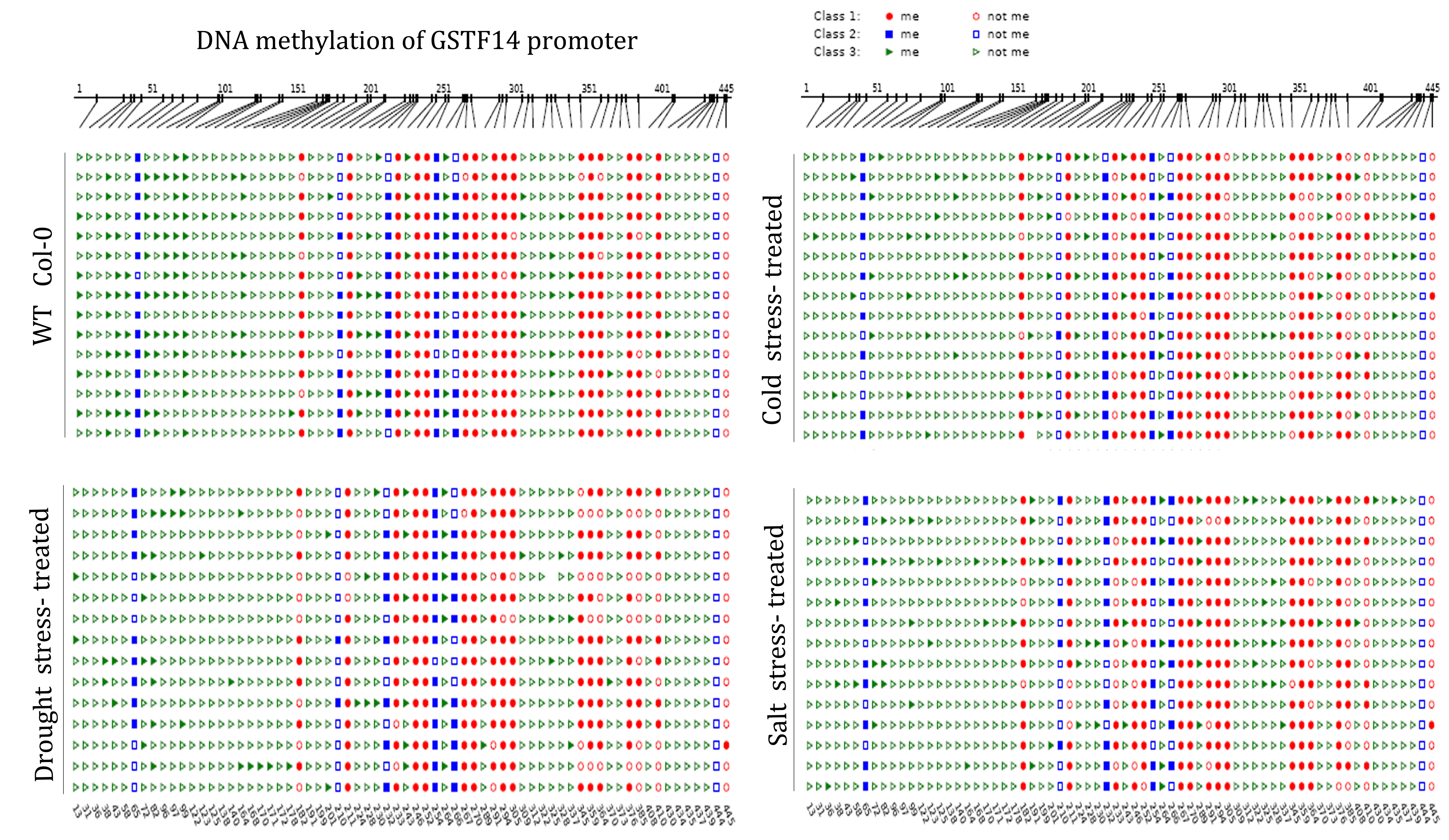
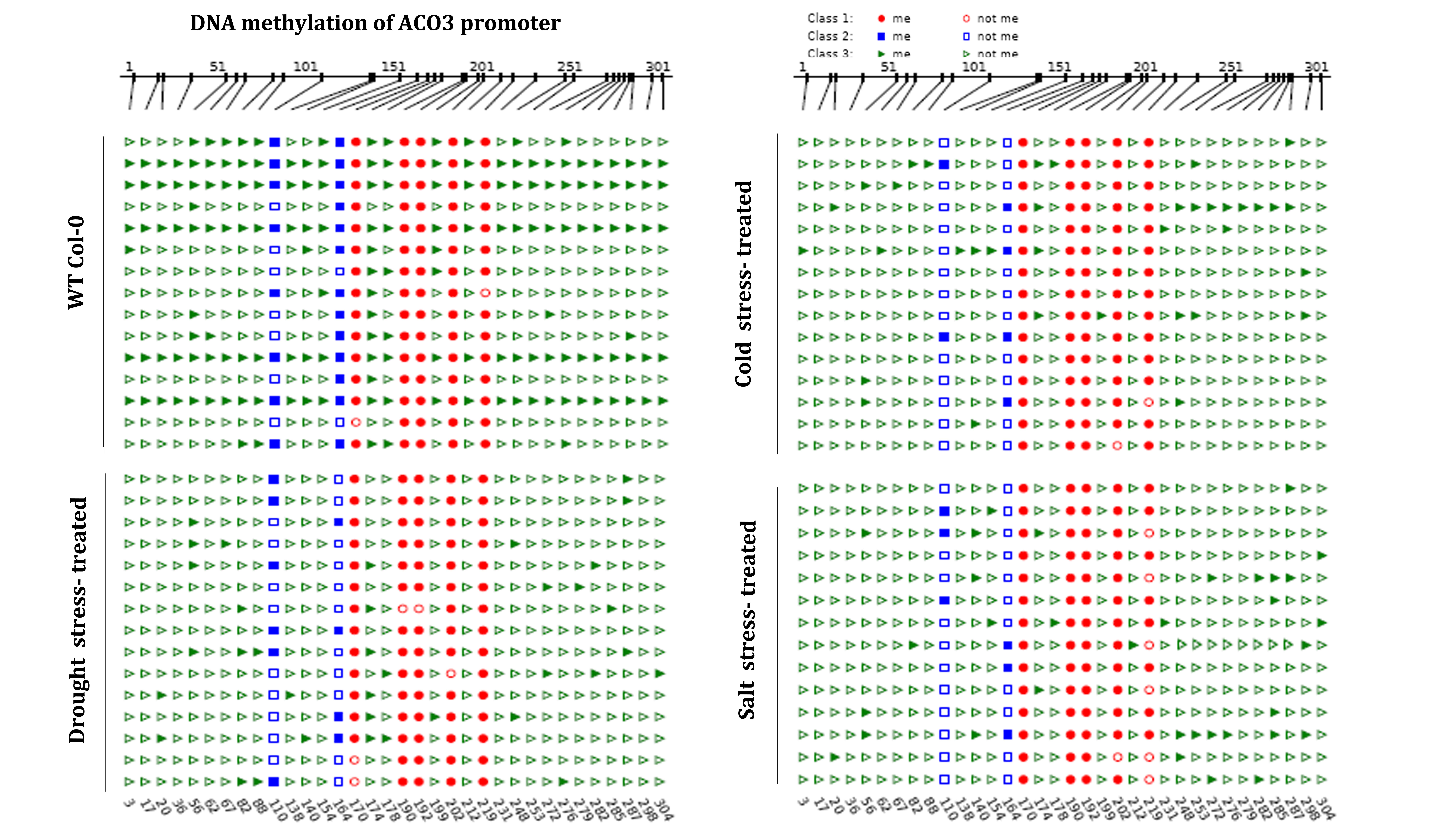
Direct correlation between the increased expression of defense and stress resistance genes and the reduction in promoter DNA methylation
To investigate whether the increase in the expression of these defense and stress resistance genes was related to the changes in their promoter DNA methylation, the DNA methylation of the plants under stress treatments was detected and compared. Untreated Arabidopsis Col-0 plants were used as the controls. After drought-stress treatment, the CG, CNG, and CHH methylation of the repeats in the ACD6 promoter decreased from 78.30% to 62.03%, from 21.67% to 8.11%, and from 13.51% to 5.80%, respectively. After cold-stress treatment, the CG, CNG, and CHH methylation of the repeats in the ACD6 promoter decreased from 78.32% to 57.77%, from 21.67% to 7.56%, and from 13.51% to 5.36%, respectively. After salt-stress treatment, the CG, CNG, and CHH methylation of the repeats in the ACD6 promoter decreased from 78.32% to 63.46%, from 21.67% to 8.26, and from 13.51% to 5.25%, respectively (Figure 3A).
Similarly, we used untreated Col-0 as a control to perform DNA methylation sequencing of the repeats in the ACO3 promoter in plants under drought-, cold-, and salt-stress treatments. After drought-stress treatment, the CG methylation of the repeats in the ACO3 promoter did not change significantly, while the CNG and CHH methylation of the repeats in the ACO3 promoter decreased significantly, from 65.89% to 33.33% and from 42.22% to 8.89%, respectively. After the cold-stress treatment, the CG methylation of the repeats in the ACO3 promoter did not change, while the CNG and CHH methylation of the repeats in the ACO3 promoter decreased significantly, from 65.89% to 20% and from 42.22% to 8.16%, respectively. After salt-stress treatment, the CG methylation of the repeats in the ACO3 promoter did not change significantly, while the CNG and CHH methylation of the repeats in the ACO3 promoter decreased significantly, from 65.89% to 21.43% and from 42.22% to 9.19%, respectively (Figure 3B).
DNA methylation of the GSTF14 promoter was analyzed next. After drought-stress treatment, the CG, CNG, and CHH methylation of the repeats in the GSTF14 promoter decreased from 90.30% to 75.49%, from 64.04% to 48.61%, and from 20.78% to 8.72%, respectively. After cold-stress treatment, the CG methylation of the repeats in the GSTF14 promoter decreased, from 90.30% to 73.03%, the CNG and CHH methylation decreased from 64.04% to 51.46% and from 20.78% to 9.63%, respectively. After salt-stress treatment, the CG methylation of the repeats in the GSTF14 promoter decreased, from 90.30% to 75.50%, the CNG and CHH methylation decreased, from 60.60% to 52.75% and from 20.78% to 8.65%, respectively (Figure 3C). Our results revealed that drought, cold, and salt stresses could induce DNA demethylation of the repeats in the gene promoters and increase the expression of these defense and stress resistance genes. Moreover, under drought, cold, and salt stresses, the pattern of DNA methylation variation of the ACD6 and GSTF14 promoters was different from that of the ACO3 promoter.
Figure 3 Analyses of DNA methylation of the promoters in plants treated with different stresses
(A) Percentage of DNA methylation in the repeat regions of the ACD6 promoter in plants treated with different stresses and untreated Col-0 plants. (B) Percentage of DNA methylation in the repeat regions of the ACO3 promoter in plants treated with different stresses and untreated Col-0 plants. (C) Percentage of DNA methylation in the repeat regions of the GSTF14 promoter in plants treated with different stresses and untreated Col-0 plants. Fifteen individual clones of each genotype were used for sequencing, and the original data are shown in supplement Data S1. The statistical analysis was performed using OriginPro 8 (http://www.originlab.com). Values are means ± SEM, and asterisks indicate statistically significant differences compared with control plants (one-way analysis of variance, P < 0.05).
Role of ROS1 in the regulation of the SA pathway in response to abiotic stresses
To further study the molecular mechanisms underlying the functioning of defense genes of the SA pathway in response to abiotic stresses, we used RNA gel blotting to detect the expression of related genes in plants mutated at key functional elements of the RdDM pathway. The results showed that the expression of ACD6 and GSTF14 clearly increased in the mutant ago4 and DNA methyltransferase mutants met1, drm1/2 and cmt3 with ecotypes Col-0 as controls (Figure 4A). RT-qPCR results further confirmed that ACD6, GSTF14, and ACO3 were upregulated in the ago4 mutant (Figure 4B), indicating that RdDM has an important role in maintaining the low transcription levels of ACD6, GSTF14, and ACO3 in wild-type plants; however, these mutants showed increased transcript levels for those genes. Repressor of silencing 1 (ROS1) can negatively regulate the RdDM pathway [8, 9]. The results further showed that the expression levels of these genes were lower in the ros1 and rdd mutants, when the Col-0 plants were used as the control (Figure 4C).
To determine whether ROS1 plays a role in the responses of these genes to abiotic stress, we performed cold-stress treatment on loss-of-function ros1 mutants and compared the expression of the ACD6 gene between the cold stress-treated ros1 mutants (ros1+cold) and the cold stress-treated Col-0 (Col-0+cold). The results showed that when Col-0 was used as the control, the expression of ACD6 in the cold stress-treated Col-0 plants significantly increased. However, the increase in ACD6 expression in the cold stress-treated ros1 mutants and loss-of-function ros1dml2dml3 (rdd) mutants was significantly inhibited when compared with the cold stress-treated Col-0 plants (Figure 4D). ROS1 plays an important role in the activation of defense and stress resistance genes in response to abiotic stress, and this finding was confirmed by the expression levels of GSTF14 and ACO3. When the cold stress-treated Col-0 plants were used as the control, the increase in GSTF14 and ACO3 expression was inhibited in the cold stress-treated ros1 mutants (Figure 4D).
Sequencing analysis confirmed that the DNA methylation levels of the repeats in the ACD6 promoter in cold stress-treated Col-0 plants were significantly reduced, including the CG, CNG and CHH sites, while the decrease in DNA methylation levels of the repeats in the ACD6 promoter in cold stress-treated ros1 mutants was obviously inhibited (Figure 4E). The results further demonstrated that the DNA methylation at CNG and CHH sites in the ACO3 promoter in cold stress-treated Col-0 plants was significantly decreased, while the decrease in DNA methylation at CNG and CHH sites in the ACO3 promoter in cold stress-treated ros1 mutants was obviously inhibited (Figure 4F).
Our results revealed that the activation of the expression of the regulator ACD6 in the SA defense pathway, the stress resistance gene GSTF14 and ABA pathway-related gene ACO3 by abiotic stresses was related to ROS1-mediated DNA demethylation.
Figure 4 Analyses of DNA methylation and the expression levels of genes
(A) Analyses of the expression levels of ACD6 and GSTF14 in the mutants ago4, met1, drm1/2 and cmt3 by northern blotting; wild-type Col-0 ecotype served as background controls for the mutant genotypes. (B) Analyses of the expression levels of ACD6, ACO3, and GSTF14 by RT-qPCR in DNA methylation mutant plants ago4, with wild-type as background control for the mutant genotypes. (C) The related genes were detected in the Col-0, ros1 and rdd mutants by RT-qPCR. (D) The related genes were detected in the untreated Col-0, the Col-0 treated with cold stress, ros1, and ros1 dml2 dml3 (rdd) mutant plants treated with cold stress by RT-qPCR. (E) Analyses of DNA methylation in the repeat regions of the ACD6 promoter in Col-0, and Col-0 plants and ros1 mutants treated with cold stress. (F) Analyses of DNA methylation in the repeat regions of the ACO3 promoter in Col-0, the Col-0 plants, and ros1 mutants treated with cold stress. The statistical analysis was performed using OriginPro 8 (http://www.originlab.com); asterisks indicate statistically significant differences compared with control plants (P < 0.05).
{kind=link}
{kind=link}
{kind=link}