Analysis of gut microbiota, microbiome and fecal metabolome during stage 1 NEC (NEC-1).
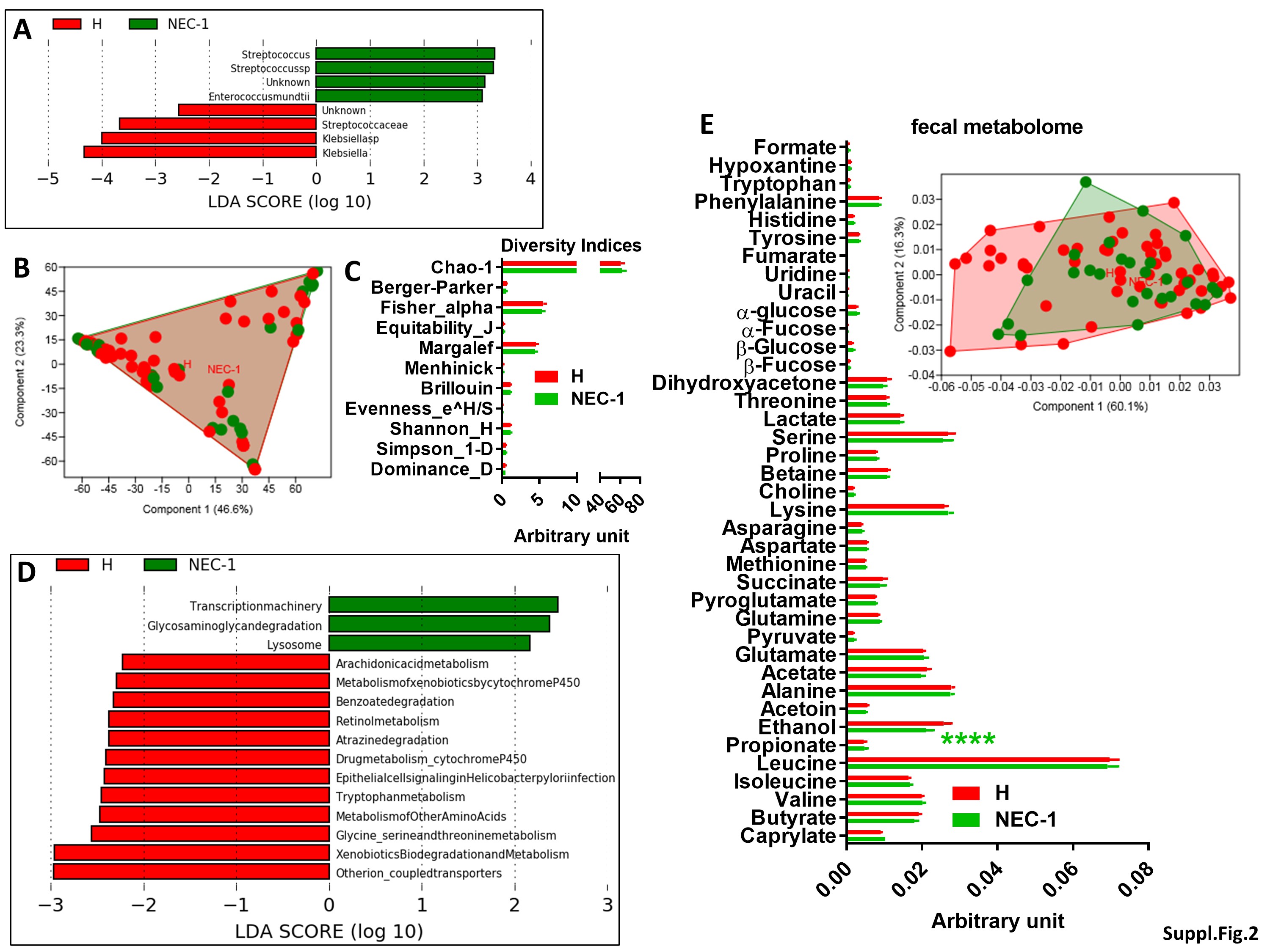
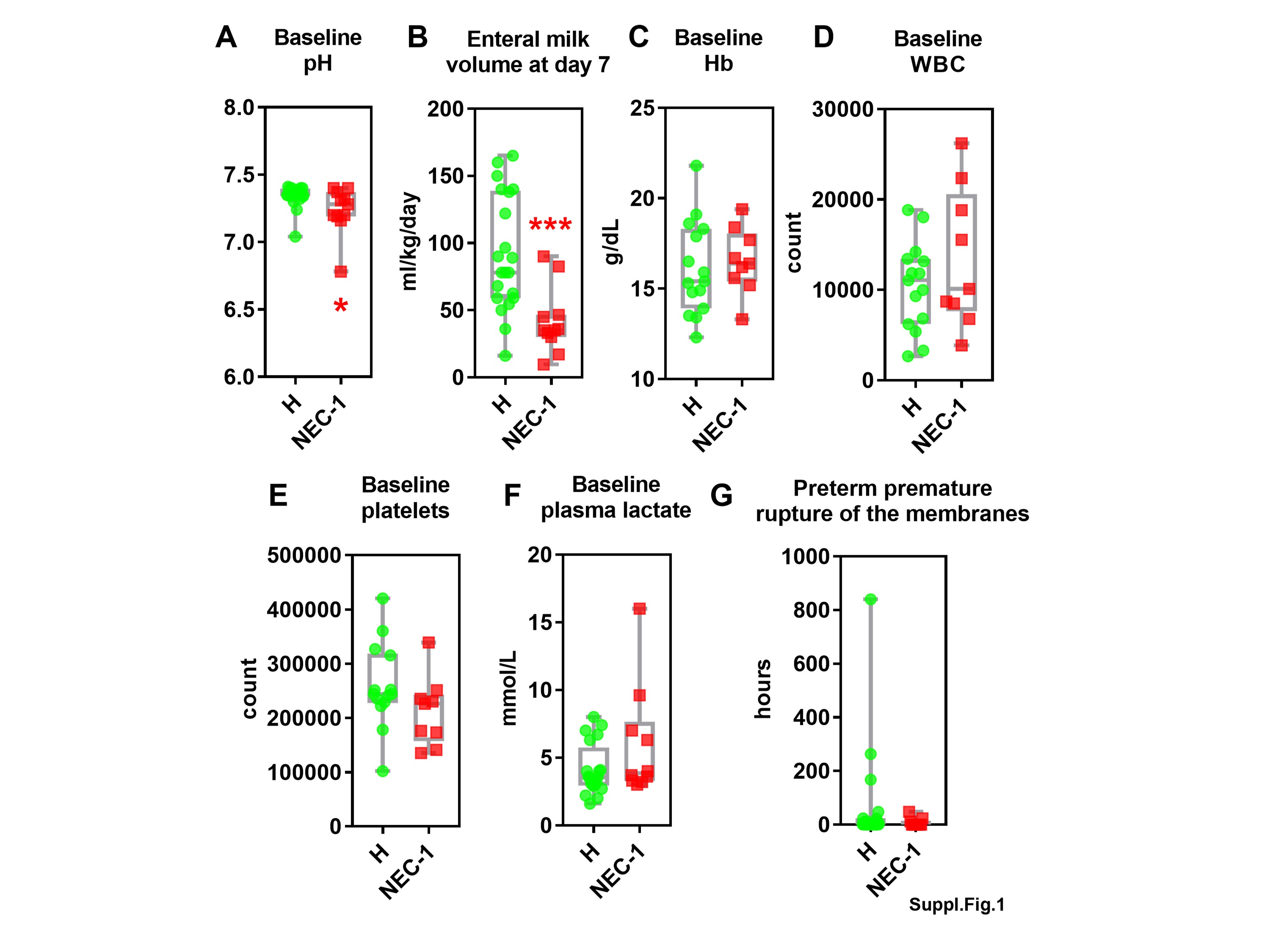
To understand the microbial and metabolomic evolution during the early onset of necrotizing enterocolitis (NEC), we studied clinical profile suspected (stage-1) NEC (NEC-1) preterm infants. NEC-1 children underwent more glycopeptides treatment, showed significantly higher cordon lactates, bacteremia and a longer full enteral feeding, when compared to age-matched healthy children (Table 1). NEC-1 children also displayed a lower plasma pH and enteral milk volume at day 7 (Suppl. Figure 1A,B) and a higher abundance of Streptoccoccus species (Suppl. Figure 2A) compared to healthy children. Both populations of children showed a high intragroup variance in terms of gut microbiota (Suppl. Figure 2B) and overall microbial diversity (Suppl. Figure 2C). NEC-1 microbiome showed increased activity for pathway related to transcription, glycosaminoglycan degradation and lysosome, compared to healthy children (Suppl. Figure 2D). Then, we analysed the fecal metabolome to appreciate NEC-1-induced changes in gut microbial metabolic activity. NEC-1 children displayed a reduced intragroup variation and significantly lower levels of ethanol (Suppl. Figure 2E). Overall, these data show that NEC-1 is characterized by a precise gut microbiota, microbiome and gut microbial metabolites profile.
Analysis of gut microbiota, microbiome and fecal metabolome during the evolution of NEC-1 over decades up to the second month of life.
Given the presence of a NEC-1-specific gut microbiota and microbiome profile, we aimed at identifying at what time these profiles establish. We divided both NEC-1 and healthy children populations in subgroups according to decades (period of ten days of life) as it follows: 1-10d (d stands for “days”), 11-20d, 21-30d for the first month of life and > 30d for the second one. In the first decade, NEC-1 children displayed a divergent and more homogenous gut microbiota compared to healthy children, with the latters characterized by a higher abundance of Klebsiella species (Fig. 1A-B). At this stage of life, gut microbiota in NEC-1 had a lower diversity based on Chao-1 index (Fig. 1C) and a different microbial activity related to replication, recombination and repair proteins, lysosome and glycosaminoglycan degradation (Fig. 1D). No significant changes were observed in fecal metabolites (Fig. 1E). Overall, these data show that gut microbiome starts to diverge at the early onset of NEC-1.
In the second decade, NEC-1 gut microbiota was characterized again by a higher abundance of Streptococcus species and bacteria from the Micrococcales order (Fig. 2A), with a high intragroup variance (Fig. 2B). At this stage of life, NEC-1 gut microbiota also showed a higher diversity based on Chao-1 index (Fig. 2C), but no microbial pathway differently regulated (Fig. 2D). As for the fecal metabolome, NEC-1 children displayed significant lower levels of serine (Fig. 2E). Overall, these data show a stronger evolution of gut microbiota than gut microbiome in the second decade, between NEC-1 and healthy children.
In the third decade of life, changes in NEC-1 gut microbiota compared to healthy children occurred to a bigger extent and were related to increased Staphylococcus and Streptococcus species (Fig. 3A-B), together with a high intragroup variance (Fig. 3C) and no change in the overall diversity indices (Fig. 3D). We also observed a NEC-1 microbiome profile mainly based on thiamine and seleno-compound metabolism (Fig. 3E). The NEC-1 gut microbiota profile of the third decade of life was associated with: i) multiple diseases and found significantly increased in ulcerative colitis (Fig. 4A); ii) host genetic variation and significantly related to ANP32E, a gene involved in ulcerative colitis [15], in line with previous reports. In terms of fecal metabolome, we observed no significant changes in NEC-1 vs. healthy children (Fig. 4C). Then, we studied feces collected in the second month of life. In this period of life, the taxonomical differences in the gut microbiota of NEC-1 vs. healthy children were related to the increase in Raoultella species in NEC-1 gut microbiota (Fig. 5A), with a still high intragroup variance (Fig. 5B) and no change in the overall microbial diversity indices (Fig. 5C). We also observed microbial functions related to DNA repair increased in the NEC-1 gut microbiome (Fig. 5D). This period of life was characterized by the highest separation in terms of fecal metabolome, with significant lower levels of ethanol and leucine in NEC-1 children.
Specific impact of NEC-1 on the evolution of gut microbiota. microbiome and fecal metabolome over the first two months of life, compared to healthy children.
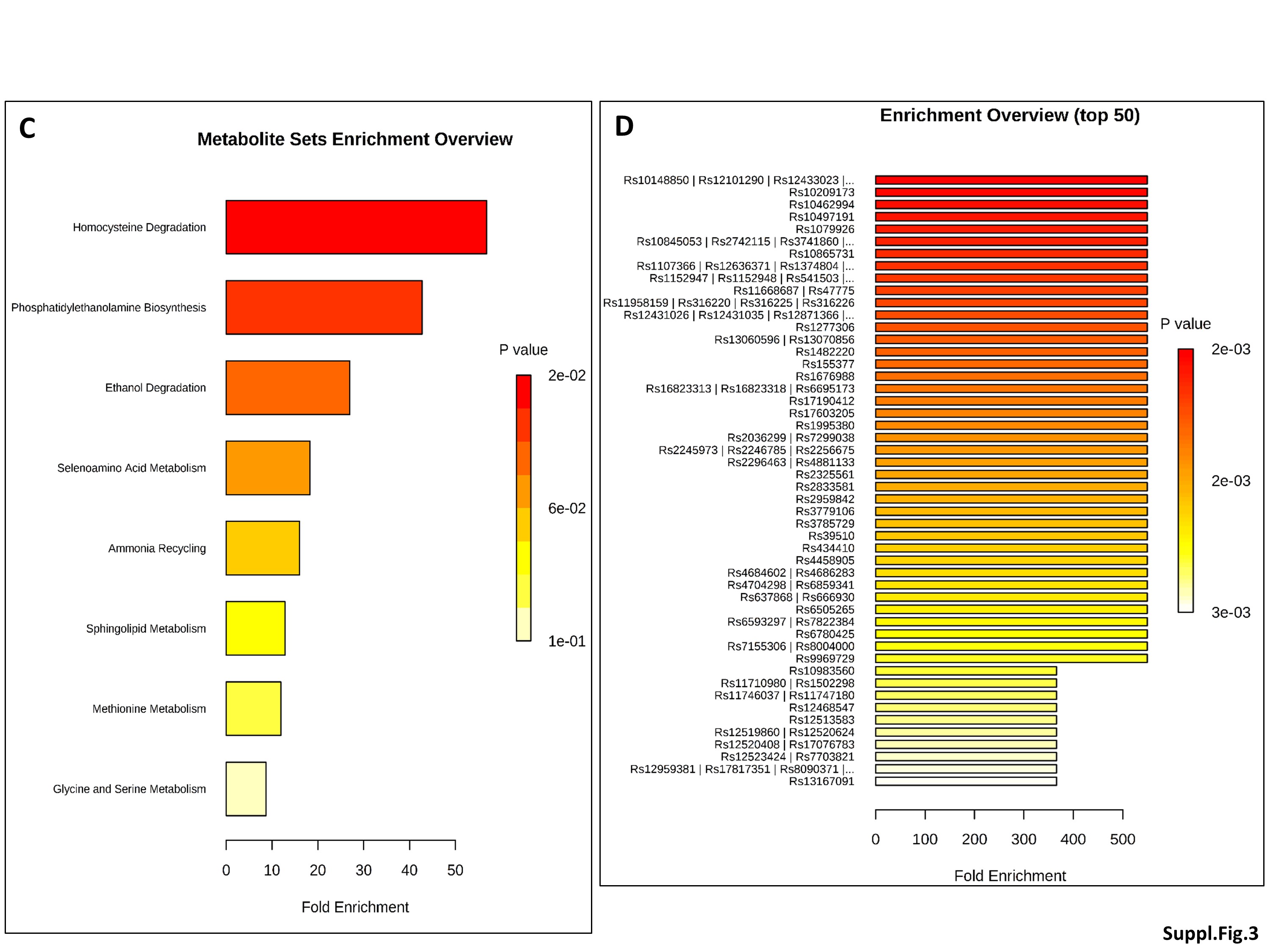
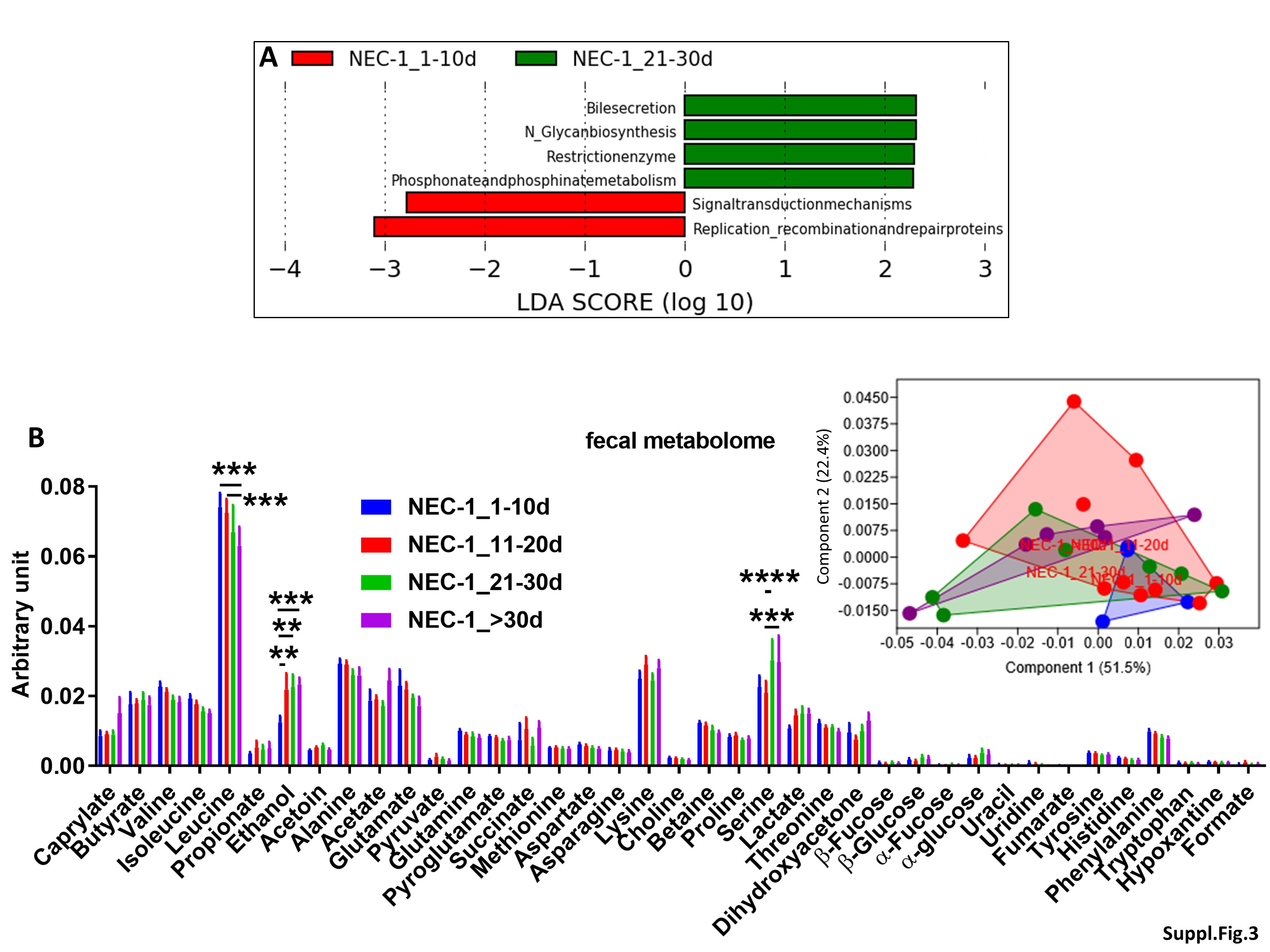
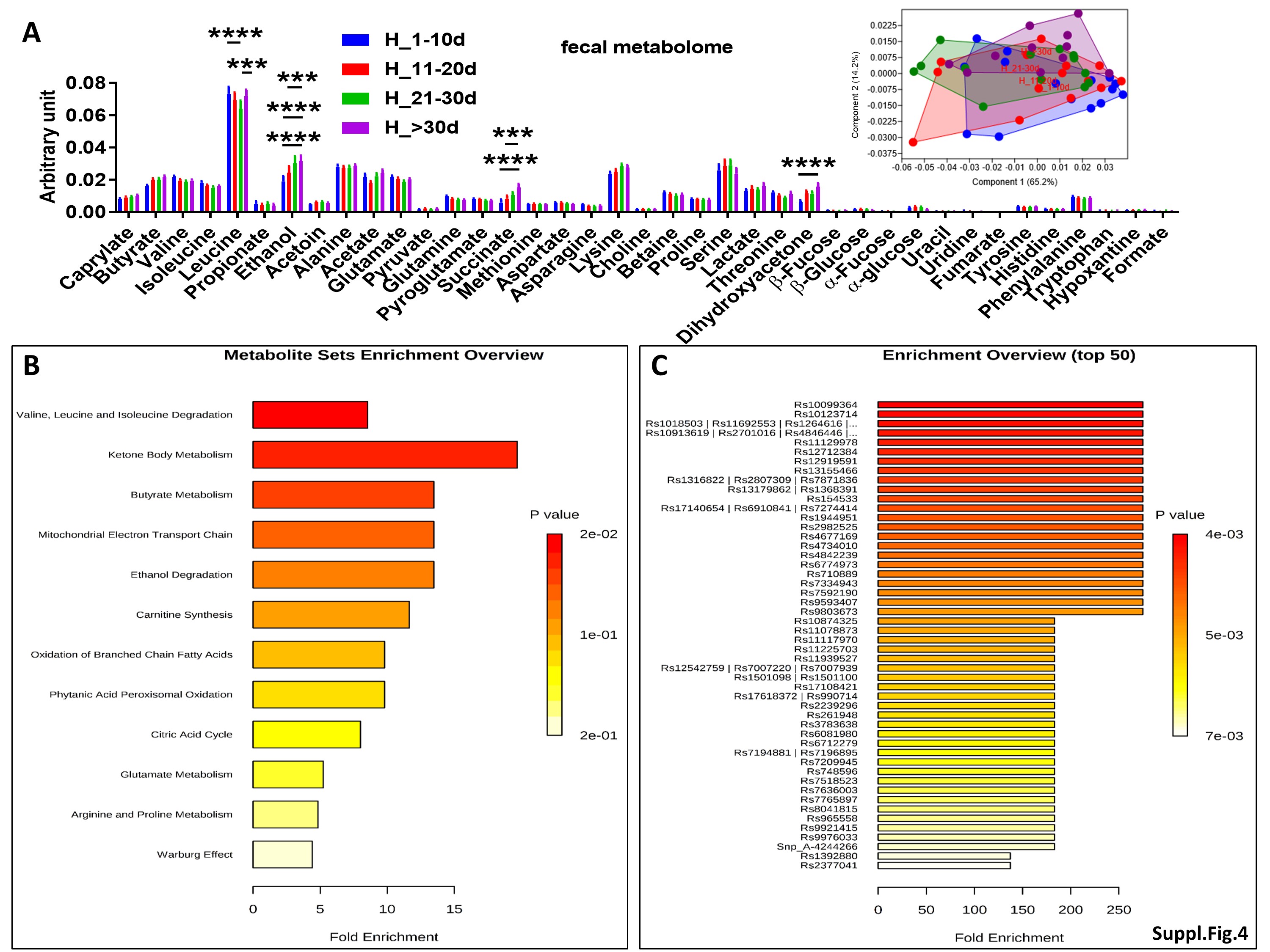
To investigate the evolution of gut microbiota, microbiome and fecal metabolome over the first two months of life, we conducted an intra-group study in both NEC-1 and healthy children, according to the four groups reported above: 1-10d, 11-20d, 21-30d and > 30d. We did not observe any taxonomic significant change in the gut microbiota of NEC-1 children. However, the group NEC-1_21-30d had a specific gut microbiome with an increased restriction enzyme activity, among others (Suppl. Figure 3A). The four NEC-1 groups also differed in terms of fecal metabolome, with regard to leucine, ethanol and serine amounts (Suppl. Figure 3B). Based on these results, we performed a metabolomic enrichment analysis on two levels: i) pathway-associated metabolite sets (Suppl. Figure 3C) and ii) single nucleotide polymorphism (SNP)-associated metabolite sets (Suppl. Figure 3D). NEC-1 metabolomic profile (increased ethanol and serine) was significantly associated to both homocysteine degradation and phosphatidylethanolamine biosynthesis (Suppl. Figure 3C), with serine being the metabolite the most linked to NEC-1-associated SNP (Suppl. Figure 3D). By contrast, in healthy children the four groups reported above did not differ in terms of both gut microbiota and microbiome, but only with regard to fecal metabolome (Suppl. Figure 4A). Healthy metabolomic profile (increased leucine, ethanol and dihydroxyacetone) was significantly associated to valine, leucine and isoleucine degradation and to ketone body metabolism (Suppl. Figure 4B), with leucine being the metabolite the most linked to healthy-associated SNP (Suppl. Figure 4C). Overall, these data suggest that: i) a different intragroup evolution exist between NEC-1 and healthy children with regard to gut microbiota and microbiome and ii) NEC-1 microbiome appears to be more sensitive to mother-related factors.
Maternal and child Factors influencing the gut microbiota, microbiome and fecal metabolome during NEC-1.
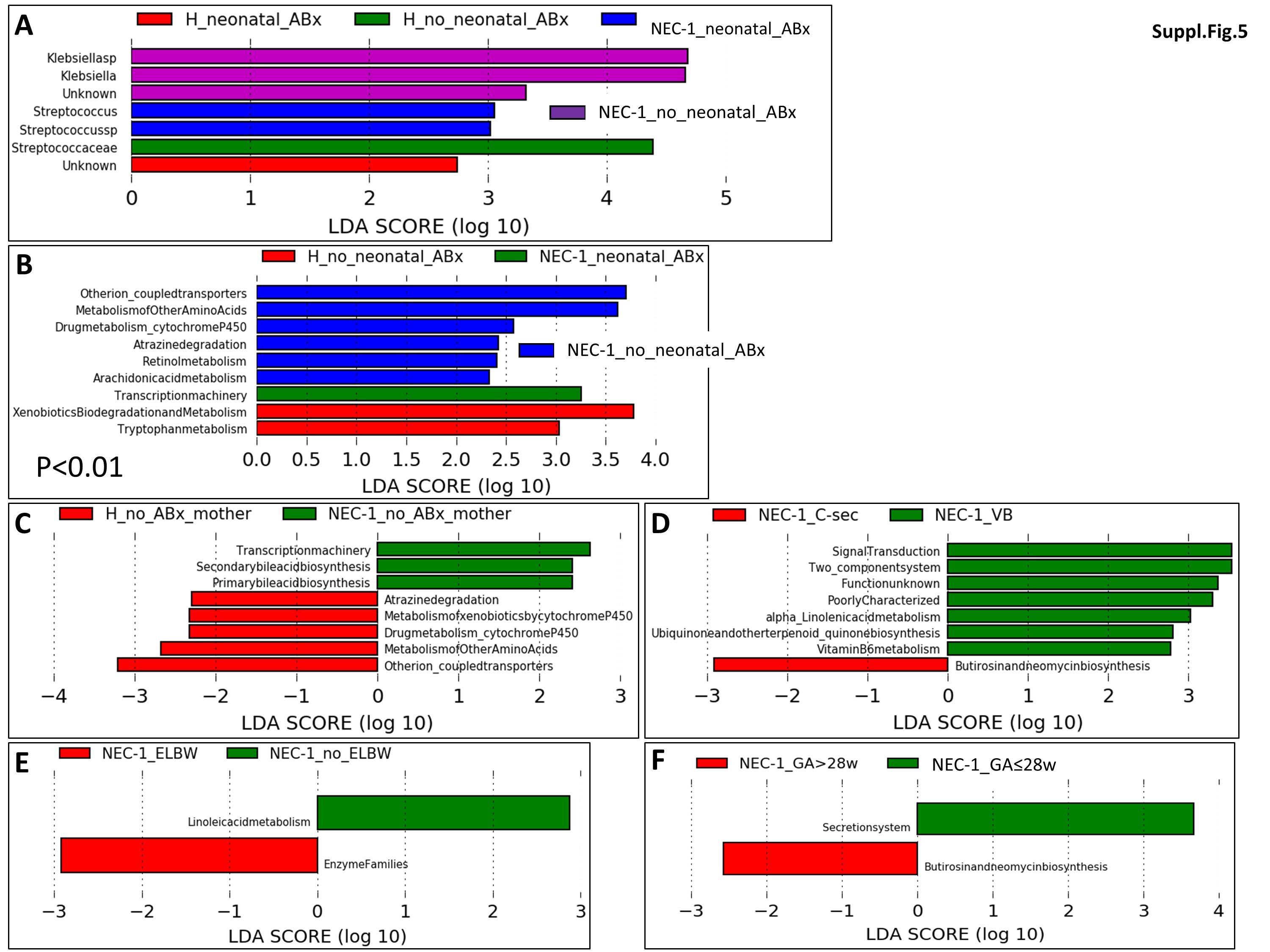
Next, we asked which factor related to both mother and child may affect the most the above reported parameters. We analysed six conditions: neonatal antibiotherapy (ABx), ABx treatment on mother, childbirth (C-section (C-sec) vs. vaginal birth (VB)), very low birth weight (VLBW), extreme low birth weight (ELBW) and the gestational age (GA) > or ≤ 28 weeks.
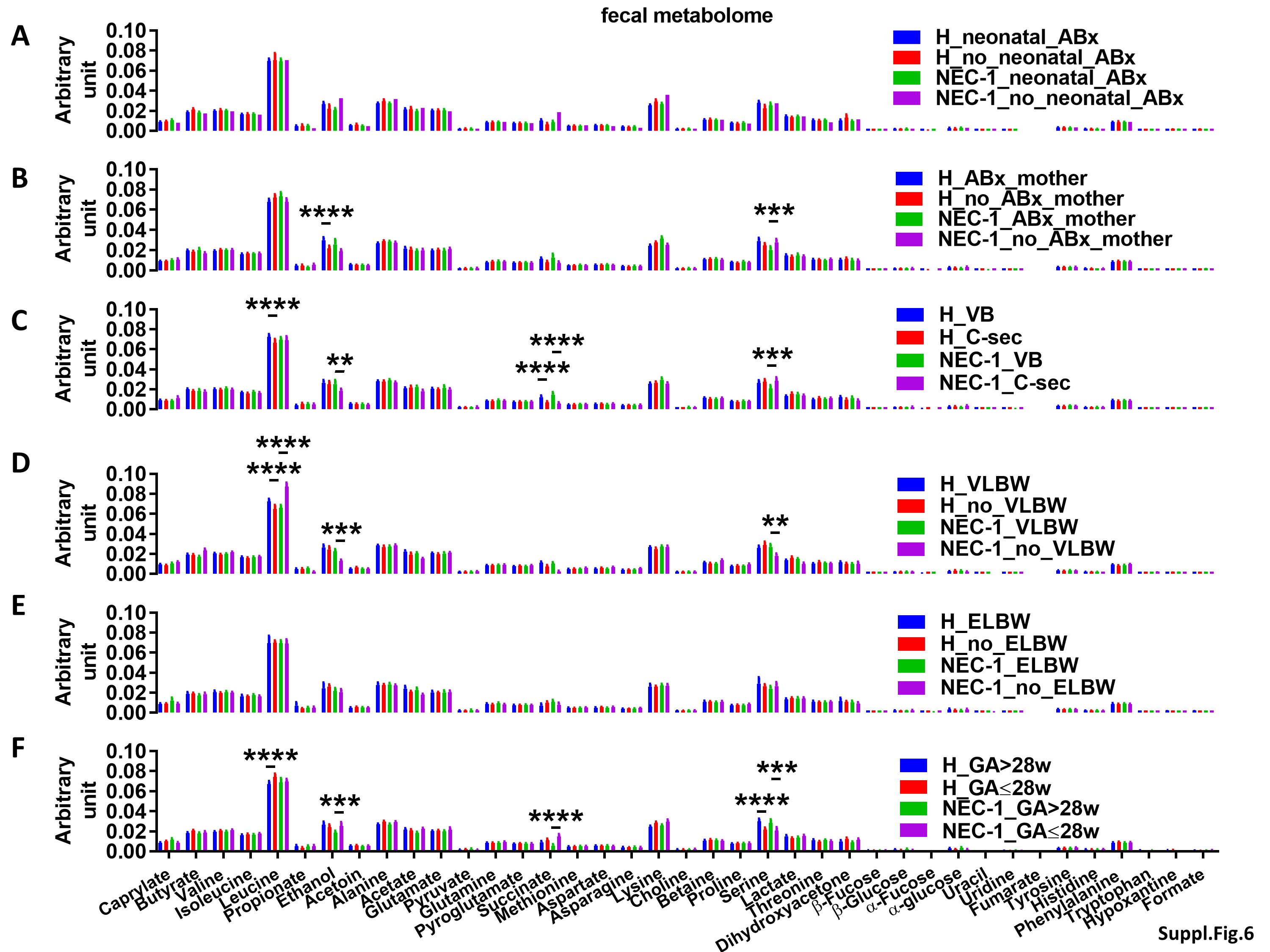
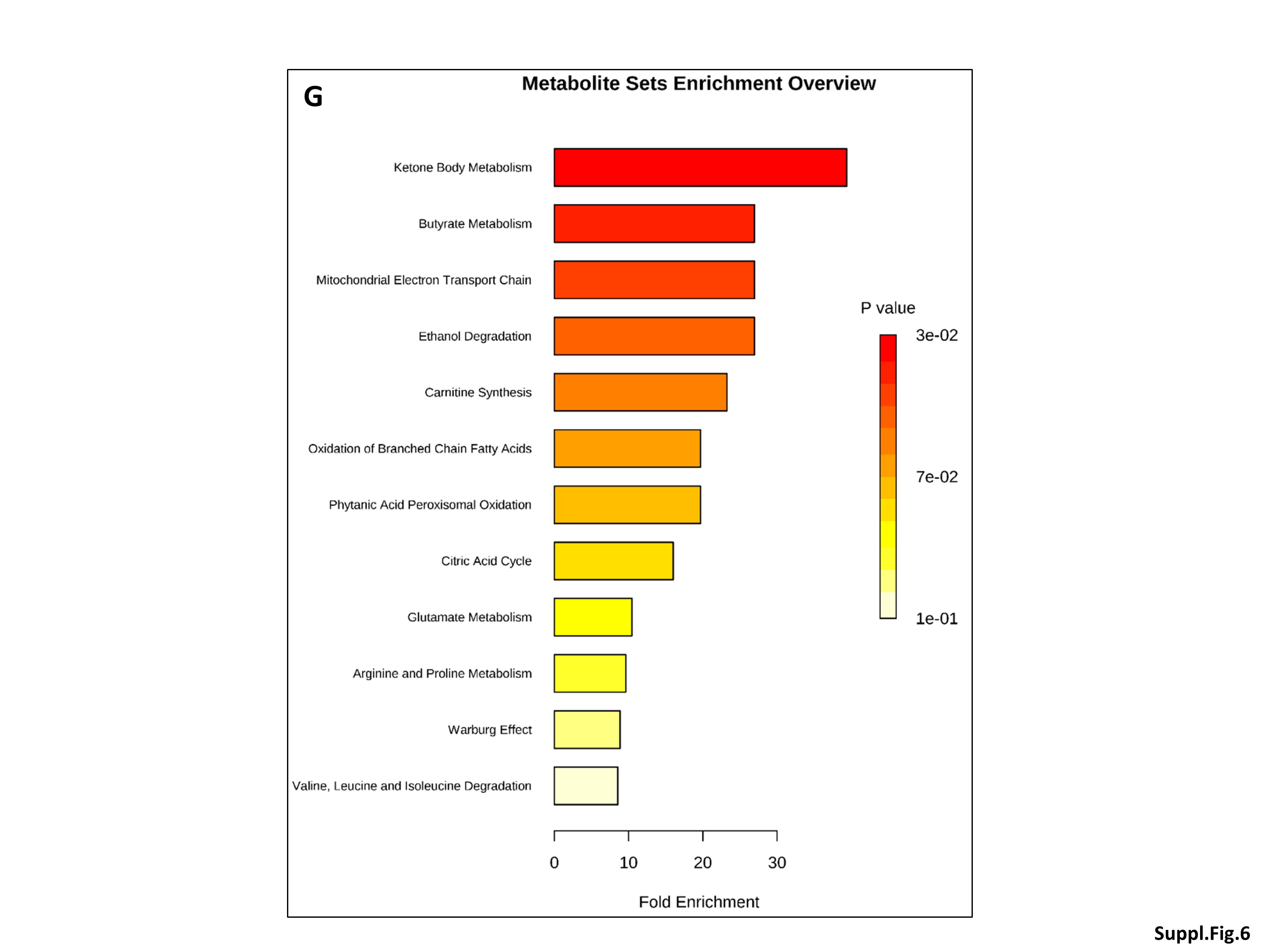
Only neonatal ABx treatment affected the gut microbiota in both NEC-1 and healthy children (Suppl. Figure 5A). By contrast, all the above factors, except the VLBW, affected the gut microbiome (Suppl. Figure 5B-F). Note that childbirth modality, ELBW and GA affected the gut microbiome only in NEC-1 children (Suppl. Figure 5D-F). Moreover, all the above factors, except the neonatal ABx treatment and ELBW, affected the fecal metabolome between NEC-1 and healthy children (Suppl. Figure 6A-F). Then, we performed again a metabolomic enrichment analysis on the pathway-associated metabolite sets, based on Suppl. Figure 6F, in which there is an increase in ethanol and succinate within in the NEC-1_GA ≤ 28w. Ketone body and butyrate metabolism were the most significantly associated with this metabolomic set (Suppl. Figure 6G).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}