The transcriptome of the early ripening berry shows strongest GLD-mediated changes
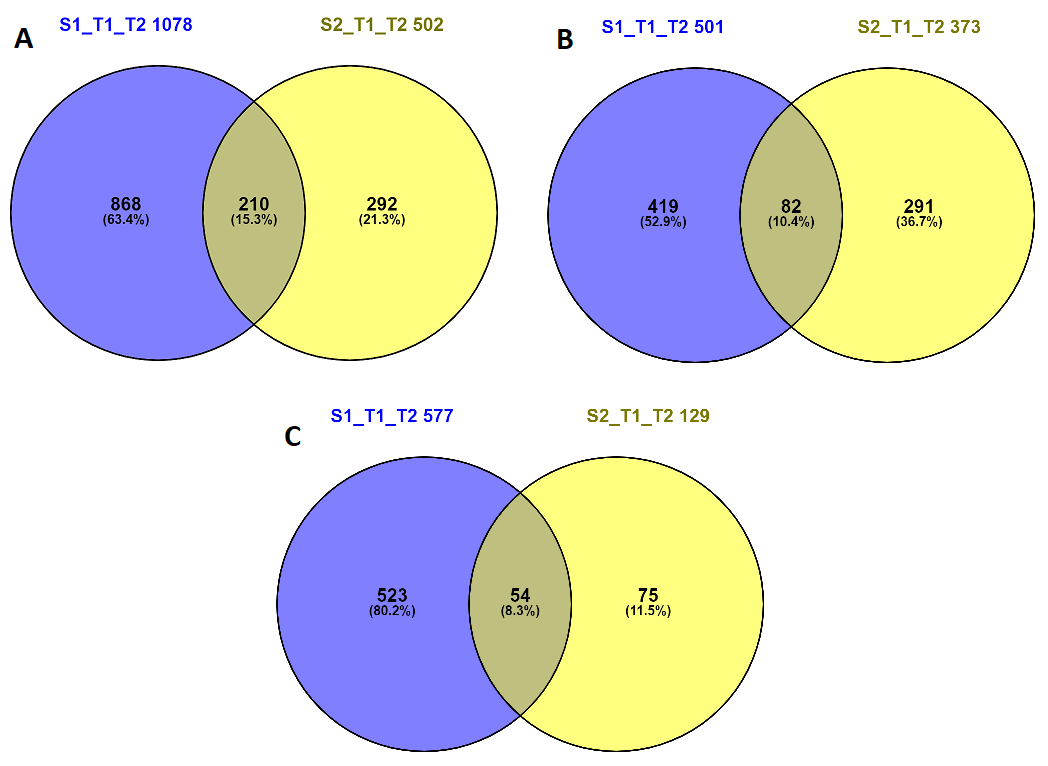
Results from PCA (Fig. 2) and Venn diagrams (Fig. 3) highlight that virus mediated gene expression is highest in the early ripening berry just after véraison. This high sensitivity of the early post-véraison berry to biotic and abiotic stresses has been already highlighted in several previous studies [21, 23, 40–42]. However, Vega et al. (2011) [19] obtained contrasted results with later ripening stage (E-L38) showing a higher number of GLRaV DEGs than at véraison (E-L35). Besides, in the latter study sampling stages are not very well defined and sampling did not account for berry heterogeneity which might be the explanation for the contradictory results.
Together these results highlighted the importance of our sampling reconstitution strategy and will prove its contribution to understand the exact triggering changes related to GLD independently of phenological delay induced by the virus.
GLD represses cell wall related transcript at véraison
Eight Xyloglucan endotransglycosylase/hydrolase (XTH) transcripts, putatively involved in cell wall extensibility and growth [43] were downregulated by T2 at S1 (Additional file 4: Table S3) with 3 amongst them (VIT_05s0062g00610, VIT_11s0052g01200, VIT_11s0052g01270) being as well repressed by T1 at S1. T. XTH regulation upon biotic and abiotic stress defers in plants with respect to applied stress. For example, Rienth et al. (2014) [21] showed an induction of XTH transcripts in green berries in response to temperature. In rice, XTHs were upregulated in the early stages of drought stress, indicating their role in the maintenance of root growth [44]. Moreover, the same specie exposed to different abiotic stresses (drought, cold, heat) exhibited an overexpression of OsXET9, a xyloglucan modifying enzyme which could represent a multiple stress-responsive gene [45]. Similarly, an increased level of transcripts for XTH gene isolated from hot pepper (Capsicum annuum) in transgenic plants of Arabidopsis and tomato corroborated the role of XTH in enhancing tolerance to drought and salt [46, 47]. However, other reports elucidated a down-regulation of XTH genes in sweet potato and Arabidopsis under cold stress [48, 49], which highlighted a contrasting behavior of these genes at different stresses [50]. As well, Maldonado-Mendoza et al. (1993) [51] and Divol et al. (2007) [52] demonstrated that expression of XTH genes can be induced during arbuscular mycorrhizal symbiosis or regulated by parasites, respectively. The here observed down-regulation of XTHs indicates a virus impact on cell wall structure and composition at early ripening berry consistently with the delayed berry growth commonly observed in virus infected berries [53, 54] and which is interestingly even observable when berry samples were reconstituted according to their sugar and acid content.
Heat shock responsive transcript are modified by GLD infection
Essential for cell growth and viability, heat shock proteins (HSPs), organized into HSP100s, HSP90s, HSP70s, HSP60s (cpn60s), and small heat-shock proteins (sHSPs) [55], function as molecular chaperones in maintaining protein quality and folding, and are stress-responsive proteins protecting plants from damage under stress [56, 57]. Under conditions of stress the increased expression and accumulation of stress proteins facilitates the ability of cells to both repair and synthesize new proteins and to replace those that were damaged as a consequence of the stress [58].
Interestingly in the present study Heat shock response related genes showed opposed expression by GLD at S1 and S2 (Table 3; Additional file 4: Table S3). At S1, heat shock related functional categories including protein folding, chaperon mediated protein folding, HSP-mediated protein folding; were upregulated by T2 and more profoundly within T1 induced genes (Additional file 4: Table S3). T1 caused an enhancement of expression of several isogenes coding for small heat-shock proteins (sHsp; HSP kDa 16.9, HSP kDa 17.4, HSP kDa 17.5, HSP kDa 17.6, HSP kDa 18.2, HSP kDa 22) among which HSP kDa 16.9 (VIT_18s0001g01610), HSP kDa 17.6 (VIT_04s0008g01610, VIT_13s0019g02760) and HSP kDa 18.2 (VIT_13s0019g02820) were the most induced.
These results highlight the important role of HSP genes in plant development and stress responses. Several biotic and abiotic stresses such as extreme temperatures, drought and pathogens are associated with an enhanced accumulation of chaperones from HSP superfamily, namely HSP70 [59–61], HSP100 (Clp protease) [62], and small HSPs [63–65], but also others such as chopper chaperone [66, 67], serpins [68–70], and protein disulfide isomerase [60, 65, 66]. Other authors observed an induction of genes belonging to this family in response to cold [71]. Furthermore, previous reports have shown that expression of HSPs is associated with the intensity of the stress that they are naturally exposed to [58, 72].
According to the literature HSP90s are considered to have a general role in the refolding of misfolded proteins that can accumulate during plant development or in response to various biotic and abiotic stress conditions [73–75], sensing the environment and mediating appropriate phenotypic plasticity [76, 77]. During the last few years, it has become evident that HSP90 is involved in diverse signal transduction pathways of disease resistance in plants due to its ability to link plant receptors to other components pivotal for triggering an appropriate defense responses against pathogens [78–80]. Here, HSP90 (VIT_16s0050g01150) expression changed in response to infection level (T1 and T2) and showed opposed expression at S1 and S2 being firstly activated and then repressed.
In parallel, HSP70s are involved in nearly every aspect of protein biogenesis during normal growth development of an organism as well as under various stresses [81–85]. However, the cellular mechanisms of HSP70 function under stress conditions are not fully understood. Plant HSP70 genes are encoded by a highly conserved multi-gene family. 14 genes encode HSP70s in Arabidopsis and there are at least 12 in spinach [86, 87]. Nicotiana and Arabidopsis transgenic lines with enhanced levels of HSP70 exhibited greater tolerance to salt, water, and high-temperature stress [88–92]. In the present study, HSP70 transcripts showed again opposed expression in response to GLD at S1 and S2. Several Hsp70 coding isogenes (VIT_02s0025g02140, VIT_05s0020g03330, VIT_06s0004g04470, VIT_08s0007g00130, VIT_11s0037g00510, VIT_18s0041g01230) were upregulated within GLD at S1, among which (VIT_06s0004g04470, VIT_08s0007g00130, VIT_11s0037g00510) were repressed at S2.
Besides, It was approved that the regulation of HSP expression is under the control of diverse heat shock TFs (HSFs) whose activity, as well, is subject to different levels of control including transcriptional, post-transcriptional, translational, and post-translation regulation [93–96]. 19 putative VvHSFs were identified among the 12X version of the grape genome [97, 98] among which 3 (VvHSFA6B, VIT_00s0179g00150; VvHSFB2B, VIT_02s0025g04170; VvHSFA2, VIT_04s0008g01110) were induced within both T1 and T2 infected berries, again only at S1. VvHSFB2A (VIT_16s0100g00720) was also found to be upregulated by T1 at S1. VvHSFA6B and VvHSFA2 were found to be repressed by both T1 and T2 at S2
As previously shown, VvHSFA2 was highly induced in tomato and Arabidopsis [99, 100] and it seems to play an important role in regulating some general stress-related genes particularly GolS1 (galactinol synthase 1) and APX2 (Ascorbate peroxidase 2) [98]. Putatively regulated by VvHSFA2, VvGolS1 [97, 98] encode galactinol synthases, and their proteins catalysis the formation of raffinose family oligosaccharides functioning as osmoprotectants in plant cells. Accumulation of this trisaccharide in grapevines has been known to occur after several types of abiotic and biotic stress [97, 101]. In the present study VvGolS1 (VIT_07s0005g01970) was activated at S1 and repressed at S2 within GLD treatments mainly by T1, representing a marked correlation with the activity of VvHSFA2 under GLD. Furthermore, a recently discovered Transcription factor, MBF1c (Multiprotein bridging factor 1c; VIT_11s0016g04080), acting upstream to salicylic acid, ethylene and trehalose, and was thought to be only involved in abiotic stress responses, was found to be upregulated here by T1 at S1 and down modulated by the same treatment at S2.
The double-facetted regulation of HSFs and HSPs along berry development, at véraison and late ripening, highlights their importance in abiotic as well as biotic stress adaptation within fruits. GLD up and down adaptive regulations exist within the heat shock family where some are induced at véraison (S1) and others were repressed at late stage of ripening (S2). This regulation of heat shock related transcripts highlights again the higher sensitivity of the early ripening berry and confirms that a co-infection of GLRaR-1 + GLRaV-3 triggers represents a higher stress level for the fruit.
GLD Modulation Of Transcription Factors (TFS)
Several families of transcription factors have been reported to be involved in plant biotic and abiotic stress [102–104]. Some transcription factors including NAC (NAM-no apical meristem, ATAF-Arabidopsis transcription activation factor, and CUC-cup-shaped cotyledon), bZIP (basic region/leucine zipper motif), WRKYs were found to be modulated by GLD. In fact, NAC TFs showed higher expression pattern within T2 infected berries at both stages and within T2 versus T1 at S2 (Table 3; Additional file 4: Table S3).
The NAC family has been characterized in a number of plant species that comprise a conserved NAM domain in the N-terminus. NAC transcription factors have been shown to participate in diverse biological processes, including hormone signaling [105–107], leaf senescence [108–111], secondary cell wall development and components [112–115] and have been reported to be involved in regulating plant development at different growth stages [116–125]. Interestingly, VvNAC60 (VIT_08s0007g07670), a master regulator of véraison [126], had higher expression levels in berries under T2 at S1. VvNAC22 (VIT_19s0027g00230) and VvNAC47 (VIT_08s0007g07670), reported to be characteristic of dormancy induction in grapevine [127], were down and up modulated by T2 at S1, respectively.
Moreover, much evidence revealed that NAC genes play important roles in plant resistance including abiotic and biotic stress responses [128]. VvNAC36 (VIT_03s0038g03410), which was identified as a powdery mildew dependent gene and may be part of defense system [129], was found to be upregulated within T2 infected berries at both stages. VvNAC90 (VIT_08s0007g02940) was also higher expressed by T2 at both stages. VvNAC90 was reported to respond to various stresses [130–132]. Therefore, it could be hypothesized that NAC domain genes are one of the key regulatory gene families involved in plant growth and defense.
Several bZIP transcription factors, which belong to one of the largest and most diverse families [133], responded to T1 treatment at S1 (Table 3; Additional file 4: Table S3). 55 bZIP genes were identified in the grapevine genome [134]. Plant bZIP proteins have been shown to regulate the integration and development of many organs and tissues, including seed maturation and germination [135], floral induction and development [136, 137], vascular development [138], embryogenesis [139] and photomorphogenesis [140]. They have also been found to be involved in responses to a variety of abiotic and biotic stimuli, such as drought [141], high salinity [142], heat stress [143], cold stress [144], pathogen infection [102, 145, 146], and hormone signaling, such as abscisic acid (ABA) [147, 148] and ethylene [149]. Two bZIP TFs, VvGBF3 (VIT_02s0025g01020 and VIT_15s0046g01440) were found to be significantly upregulated within T1 infected berries at S1. VvGBF3 (VIT_02s0025g01020) was reported to be activated within grapevine roots and cell cultures in response to ABA, a well described phytohormone that is functionally involved in regulating various developmental processes and stress responses [150].
Similarly, WRKYs (VIT_07s0031g01710, VIT_08s0058g01390, VIT_12s0055g00340, VIT_13s0067g03140) positively responded to GLD (Additional file 4: Table S3). Two transcripts coding for the plant-specific transcription factor WRKY70 (VIT_08s0058g01390 and VIT_13s0067g03140) were transiently upregulated by T2 at S1 or S2. WRKY70 is a common component in salicylic acid (SA)- and jasmonic acid (JA)-mediated signal pathways and was shown to be involved in the crosstalk between SA and JA-dependent defense signaling in plants [151]. Furthermore, other studies indicate that WRKY70 has a pivotal role in the balance between SA-dependent and JA-dependent defense pathways [152].
Besides, the expression profiles of VvWRKY genes in response of grapevine to GLRaV-3 infection in both véraison and ripening phases revealed a general repression of many members of the VvWRKY family, especially WRKY51 [19]. On the other hand, a WRKY9 transcript (VIT_12s0055g00340) showing higher expression level within T2 infected berries compared to T1 ones at S2. This TF was shown to be phosphorylated by mitogen activated protein kinases (MAPKs) to control plant defense signals via regulating the expression of NADPH oxidase, which triggers a ROS (reactive oxygen species) burst and cell death in Nicotiana benthamiana [153].
Effector-triggered immunity (ETI): intracellular immune receptor R (resistance protein) in response to GLD
Plants use nucleotide-binding leucine-rich repeat (NB-LRR) domain-containing resistance (R) proteins, which recognize, directly or indirectly, viral effectors (called avirulence (Avr) factors) and activate ETI in a defense mechanism similar to that employed in non-viral infections [154]. Accordingly, a Leucine-rich repeat family protein (LRR; VIT_10s0092g00570) was found to be upregulated by T2 at S1 (Additional file 5: Table S4) similar to results reported by Vega et al. (2011) [19], where this protein was upregulated by GLRaV-3 in ripening berries. Many R genes confer resistance to diverse plant pathogens including bacteria, fungi, oomycetes, insects, and viruses [155, 156]. Drawing upon previous studies, it was well established that only few R proteins show direct interactions with Avr factors [157, 158] but most of them seem to take action indirectly by means of other intermediate host proteins. Martelli (2014) indicated that no direct resistance gene interaction with Avr factors has been described within Vitis genus [159]. In the present study, several putative R genes were up-modulated mainly by T2 at S1 or S2 and downregulated by T1 at S1 (Additional file 5: Table S4) suggesting a more pronounced defense by T2. This response is in concordance with the systemic infection caused by GLRaVs in vines.
Nonsense-mediated decay pathway and RNA Silencing machinery triggered by GLD
The Nonsense-mediated decay (NMD) pathway responsible for degrading transcripts with premature termination codons (PTCs) that normally arise through improper splicing is considered as a general viral restriction mechanism in plants [160]. In addition, the RNA silencing pathway or RNA interference (RNAi) is a well-established natural antiviral defense mechanism in plants, in which the viruses are both inducers and targets of RNA silencing [161, 162]. The current state of knowledge is that an apparent and evident spatial and functional overlaps exist between the two pathways in plant [163].
In the present study, several genes with putative functions in the post-transcriptional control of berry ripening were modulated by T2 (Table 3; Additional file 5: Table S4). Five homologs of Regulator of nonsense transcripts-1 (UPF1; VIT_01s0010g01830, VIT_01s0010g01850, VIT_16s0050g02320, VIT_16s0050g02330, VIT_16s0050g02340), which has potential roles in the elimination of nonsense-containing RNAs showed higher expression levels in T2 at S1 concomitantly with genes involved in gene silencing by miRN. Plant Dicer-like (DCL) enzymes process viral RNAs into small interfering (si)RNA to trigger plant endonuclease Argonautes which repress viral gene expression [164]. DCL2 and DCL4 have thus a crucial role in viral defense mecanism [165–167]. AGO1, AGO2, AGO4, AGO5, AGO7 and AGO10 have been shown to display antiviral activity in Arabidopsis, while AGO1 and AGO18 play antiviral defense roles in rice [168]. Here, Two Argonaute proteins AGO1 (VIT_01s0010g01240, VIT_06s0061g01040) and AGO2 (VIT_10s0042g01150, VIT_10s0042g01180) were induced by T2 at S1. AGO2 was found to be upregulated also by T1 at S1. Moreover, it can be noticed that a DCL2 (VIT_04s0023g00920) showed an enhancement in T1 infected-berries only at S1. Thus, the adaptation of RNA Silencing machinery seems to be activated in véraison, but not in late ripening berries upon GLD infection. In contrast, a previous study on leafroll indicated that the viral infection caused up-regulation of DCL1 in ripening berries, which could again putatively explained by the different berry sampling strategies [19]. The antiviral RNAi response is effective in various species [169], even though it is slow and thus viral infections are often not completely cleared. Nakahara and Masuta (2014) [170] suggest that it is plausible that both RNAi and R gene-mediated pathways undergo cross-talk to maximize the efficiency of defense responses against viral infections. Together these results highlight the more pronounced defense response by the coinfection with GLRaV-1&3 (T2) as compared to T1 (GLRaV-1) and again the higher sensitivity and reactivity of the early ripening berry to biotic stressors.
Grapevine innate plant defense mechanism controlled by hormonal signaling face to GLD
Plant responses to a wide range of biotic and abiotic stresses can be mediated by defense-related mechanisms which are tightly regulated by hormone-mediated signaling pathways, mainly jasmonic acid (JA), salicylic acid (SA) and ethylene (ET) [171–173].
Jasmonic Acid
The role of JA signaling in virus defense is controversial. Several transcripts involved in the JA pathway were impacted by GLD in the present study (Additional file 5: Table S4). A Lipoxygenase protein (LOX1; VIT_06s0004g01480), the key enzyme of the JA pathway [174], was upregulated by T2 in S2. Similarly, Vega et al. (2011) indicated that LOX2 protein showed increase expression in GLRaV-3 infected-berries at ripening (E-L38) [19]. Concomitantly, Allene Oxyde Synthase (AOS; VIT_18s0001g11630), ensuring the first committed step of JA biosynthesis by converts 13-HPOT to a highly reactive allene oxide, which in the second committed step is converted to cis-12-oxo phytodienoic acid (OPDA) by allene oxide cyclase (AOC) [175], was induced as well by T2 at S1. Furthermore, other regulator genes affecting JA pathway, putatively mediated by JA-signaling, were highly upregulated in T2 infected plants at S2, such as for example Enhanced Disease Susceptibility 1 protein (EDS1; VIT_17s0000g07560), named as well Non-expressor of Pathogenesis-Related Genes (NPR1), Non-inducible Immunity 1 (NIM1) or Salicylic Acid Insensitive 1 (SAI1). NPR1 is considered as a master redox sensitive transcriptional regulator of SA response and a mediator of SA-JA crosstalk [176]. It plays a significant role in the establishment of systemic acquired resistance (SAR) and induced systemic resistance (ISR) [177] and intervenes in the regulation of Pathogenesis Related (PR) genes expression [176, 178, 179]. Consequently, several PR proteins 1 (PR-1) transcripts (VIT_18s0001g15640, VIT_18s0001g15650) were induced by T2.
Ethylene biosynthesis repression and Ethylene-mediated signaling adaptation
Ethylene is a well-known plant hormone which plays multiple regulatory roles in plant growth and development and is also involved in the response to various biotic and abiotic stress [180–183].
The biosynthesis of ethylene starts with the conversion of the amino acid methionine to S-adenosyl-methionine (AdoMet or SAM) by the enzyme AdoMet synthetase (AdoMetS) [184]. AdoMet is further converted to 1-aminocyclopropane-1-carboxylic acid (ACC) by ACC synthase (ACS), the first committed and in most cases the rate-limiting step in ethylene biosynthesis. The second step corresponds to the oxidation of ACC by ACC oxidase (ACO) to form ethylene, CO2, and hydrogen cyanide. Several studies indicate that variation in endogenous ACC levels (thus in ACS activity) strongly correlates with ethylene production rate in numerous plant tissues [184, 185], highlighting that ACS might be considered as a prime target for multiple regulatory inputs [186]. Upon GLD infection ACS (VIT_15s0046g02220) was repressed by T2 at S2 (Additional file 5: Table S4), suggesting a repression of ethylene biosynthesis. Moreover, two ACO transcripts (VIT_00s2086g00010, VIT_11s0016g02380) were found to be repressed by T2 at S1, pointing in the same direction. Similar results were reported by Blanco-Ulate et al. (2017) [27], indicating that the transcriptional networks of hormones involved in grape berry ripening, especially ABA and ethylene, were inhibited by GRBaV (Grapevine red blotch virus) infections. Interestingly and especially with regards to genes implicated in defense response and upregulated by JA, our observations are in concordance with the, in the literature described antagonism, between ET and JA in plants under biotic and abiotic stress [187]. JA and ET were shown to act antagonistically in regulating expression of wound-responsive genes [188, 189] and metabolite biosynthetic genes [190] and JA positively regulates plant defense against insect attack [191, 192], while ET functions oppositely [193–195].
Calcium signatures and signaling events modulated by GLD
Other signaling elements that are implicated in plant stress responses and especially during immune responses to pathogens, Ca2+ ions are among the earliest actors that coordinate plant adaptive responses [196, 197].
In plants, Ca2+ sensors or Ca2+ binding proteins are divided into three main classes including, calmodulin (CaM) and calmodulin-like proteins (CMLs), Ca2+-dependent protein kinases (CDPKs) and finally calcineurin B-Like proteins (CBLs) [198–200]. CaM37 ( VIT_18s0122g00180), a regulator of JA signaling [201] and known to stimulate plant defense mechanisms [202], was induced by T1 at S1 and another CaM transcript (VIT_04s0023g01100) by T2 in both stages (Additional file 5: Table S4). Furthermore a calcium-binding protein (CML: VIT_01s0026g02590), involved in calcium signaling and implicated in a variety of plants’ responses to pathogen and herbivore attack including grapevine [203, 204], was highly upregulated in T2 and less in T1 at S1 and S2. Similarly, two transcripts encoding CaMBP (VIT_17s0000g03370, VIT_17s0000g03390) implicated in plant-virus interaction, were up-regulated by T2 at both stages (Additional file 5: Table S4).
CaMBPs with functions during plant immunity have been reported [205, 206]. For example, upon infection with pathogens, various CaM-binding TFs have been implicated in plant defense responses via acting on homeostasis regulation by SA [199, 207]. Furthermore, several Ca2+-ATPases (ACA8: VIT_00s0463g00030, VIT_14s0030g02110; ACA13: VIT_05s0020g04260, VIT_05s0020g04270), which were identified as important components of receptor-mediated signaling in plant immune responses and development [208], were highly overexpressed by T2 at S1. Two out of them (VIT_05s0020g04260, VIT_14s0030g02110) were upregulated by T2 at S2, as well (Additional file 5: Table S4).
Together these expression pattern indicate that GLD infections triggers different autodefense mechanism in grapevine berries and highlight that the response is significantly more pronounced when vines were infected with both virus particles (GLRaV-1 and GLRaV-3)
GLD infection does not directly repress anthocyanin synthesis
Phenolic compounds are one of the main quality determining compounds of wines via their contribution to the organoleptic perception, particularly color and astringency and are thought to be responsible for various health benefits putatively associated with moderate wine consumption [209–212]. They principally consist of flavonoids which include flavonols, anthocyanidins and proanthocyanidins. Phenolic compounds derive from the phenylpropanoid pathway which was shown to be repressed in previous studies on berries from virus infected vines [19, 27]. In the present study we could not confirm most of the previously reported expression results of phenylpropanoid and further downstream branches. Only two isoenzymes of phenylalanine ammonia-lyase (PAL1: VIT_08s0040g01710, PAL2: VIT_13s0019g04460), the first committed step of the phenylpropanoid pathway [213] were found slightly but not significantly repressed by T1 and T2 at both stages S1 and S2 (Additional file 6: Table S5).
Further downstream of PAL, cinnamate 4-hydroxylase (C4H) catalyzes the conversion of cinnamate into p-coumarate and then 4-coumarate CoA ligase (4CL) ensures the formation of p-coumaroyl CoA. In the present study some C4H and 4CL transcripts showed only a non-significant downregulation tendency (VIT_06s0004g08150, VIT_11s0052g01090, VIT_17s0000g01790) and only one C4H transcript (VIT_11s0065g00350) was differentially repressed by T1 and T2 at S2 (Additional file 1: Table S1).
Concomitantly further downstream Chalcone synthase transcripts (CHS1; VIT_14s0068g00920, CHS3; VIT_05s0136g00260 ) encoding the first committed enzyme in flavonoid biosynthesis which catalyzes the polyketide condensation reaction of p-coumaroyl-CoA and three acetate units from malonyl-CoA to yield chalcone [214], were non-significantly modulated by both GLD treatments contrarily to Vega et al. (2011) who found CHS1 up-regulated in GLRaV-3 infected berries around véraison (E-L35 - E-L36), whereas CHS3 was down modulated at E-L35 and induced at E-L36. A down modulation of these two enzymes was detected at ripening stage (E-L38) [19].
Moreover, flavanone 3-hydroxylase transcripts (F3H1: VIT_04s0023g03370, F3H2: VIT_18s0001g14310) as well as flavonoid 3′-hydroxylase (F3′H: VIT_17s0000g07200, VIT_17s0000g07210) and flavonoid 3′5′-hydroxylase (F3′5′H: VIT_06s0009g02810, VIT_06s0009g02830, VIT_06s0009g02840, VIT_06s0009g02880, VIT_06s0009g02970, VIT_06s0009g03000, VIT_06s0009g03010), which mediate the addition of hydroxyl groups to the B ring of flavones, dihydroflavonols, and flavonols [215], were not significantly modulated by GLD treatments (Fig. 5; Additional file 6: Table S5) similarly to dihydroflavonol 4-reductase (DFR: VIT_16s0039g02350, VIT_18s0001g12800) and Leucoanthocyanidin dioxygenase (LDOX: VIT_02s0025g04720).
Otherwise, proanthocyanidins (PAs), also called condensed tannins, are synthetized through phenylpropanoid and flavonoid pathways. In grapes, PAs are under different forms (monomers, oligomers and even polymers) consisting mainly of (+)-catechin, (−)-epicatechin, (−)-epigallocatechin and (−)-epicatechin 3-gallate. PA amount and composition vary between berry compartments; In fact, PAs accumulated greatly in berry skin and seed tissues [216, 217] whereas, they are weakly present in the mesocarp [218]. One of the main regulators of PA synthesis, the TF MYBPA1 (VIT_15s0046g00170) [34, 219, 220] was found to be differentially repressed by T1 and more by T2 but only at S1 (Fig. 5; Additional file 1: Table S1; Additional file 6: Table S5). Concomitantly, the MYBPA1 regulated transcripts of leucoanthocyanidin reductase (LAR1: VIT_01s0011g02960; LAR2: VIT_17s0000g04150) and anthocyanidin reductase (ANR: VIT_00s0361g00040) [221] which are responsible for producing the flavan-3-ol monomers required for the formation of PA polymers [220] and were found to be impaired by GLD at S1. Thus, it can be hypothesized that a partial repression of early phenylpropanoid synthesis notably related to proanthocyanidin synthesis, could be detected as seen for MYBPA1, LAR1, LAR2 and ANR which were found affected by GLD only at the first stage (S1), which is congruent with previous results obtained by Vega et al. (2011) [19].
Further downstream of proanthocyanidins, vacuolar anthocyanins are generated after cytosolic modifications of anthocyanidin aglycones by glycosylation, methylation and acylation. These modifications were shown to have a crucial role in the stabilization of anthocyanin pigments [222–225] In particular, glycosylation enhances the hydrophilicity and stability of anthocyanins and is also necessary for color stability [226, 227]. A large family of glycosyltransferases (GT) ensure the O-glycosylation of anthocyanidins or anthocyanins in plants, among which the UDP-glucose: flavonoid-3-O-glycosyltransferase (UFGT) catalyzing the O-glycosylation of anthocyanidins at C3 position in V. vinifera grapes [228, 229]. Here, the UFGT transcript (VIT_16s0039g02230) was revealed not impacted by T1 and T2 (Fig. 5; Additional file 6: Table S5). Neither was one of the main regulators of anthocyanin synthesis in grapes [230, 231] VvMybA1 (VIT_02s0033g00410, Fig. 5; Additional file 6: Table S5). Together these results are partly opposed to results observed by Vega et al. (2011) [19], where UFGT1 and MYBA1 were strongly repressed by GLRaV-3 at ripening (E-L38), however MYBPA1 was also repressed by the virus infection at ripening stage (E-L38).
The non-impairment of anthocyanin synthesis by GLD infection is emphasized by the expression pattern of other important anthocyanins synthesis related transcripts such as O-methyltransferases (OMTs), acyltransferases (ACTs), glutathione S-transferase and VvanthoMATEs which were not modulated by GLD in the present study (Additional file 6: Table S5). Only one transcript coding for VvOMT2 (VIT_12s0059g01750) was differentially expressed within both GLD infected grapevines being markedly repressed mainly by T1 at S1 (Additional file 1: Table S1).
Together these results point out the non-significant impairment of anthocyanin biosynthesis related key genes such as MYBA1 and UFGT by GLD. This indicates that previously effects on anthocyanin biosynthesis caused by GLD infection, reported in other studies [19], are putatively due to a virus induced delay in phenological of berries sampled at the same time point, but not at the same ripening stage.
Sugar metabolism and transport were not directly impeded by GLD infection
Sugar accumulation starts at véraison together with fruit softening followed by the onset of anthocyanin synthesis [28]. It is well established that the accumulation of hexose sugars (glucose and fructose) in the berries implicates the activity of sucrose-metabolizing enzymes, sucrose transporters, and monosaccharide transporters (MSTs) [232].
In the grapevine genome, a small gene family with three members encodes for sucrose transporters in grapevine berries (VvSUC11/SUT1, VvSUC12, VvSUC27). The increase of VvSUC11 and VvSUC12 expression closely correlates with post-véraison sugar accumulation during ripening, unlike VvSUC27 whose quantity of transcripts significantly decrease at this same stage [233]. This suggests that VvSUC11 and VvSUC12 may have a role in the import of sucrose into the ripening berry cells [232].
In the study of Vega et al. (2011), transcript profiling showed that in GLRaV-3 infected vines the accumulation of glucose and fructose was decreased during ripening and that the expression of genes involved in sugar metabolism and transport was influenced by the virus, particularly, VvHT1 and VvMSA which underwent a significant downregulation at most developmental stages studied [19].
A different scenario is shown here where no significant modulation of the expression of the tree sucrose transporters by GLD was observed at both stages. In fact, VvSUC11 (VIT_18s0001g08210) and VvSUC27 (VIT_18s0076g00250) were slightly but non-significantly upregulated by GLD at S1 and only slightly repressed at S2. Whereas, VvSUC12 (VIT_01s0026g01960) was found barely downregulated at S1 and induced at S2. Moreover, none of the 4 cytoplasmatic neutral invertases (nINV; VIT_03s0038g01480, VIT_05s0077g00510, VIT_13s0074g00720 and VIT_18s0072g01040) cleaving sucrose into glucose and fructose, was found significantly impacted (Fig. 6; Additional file 7: Table S6). In addition, five sucrose synthases (SuSy: VIT_04s0079g00230, VIT_05s0077g01930, VIT_07s0005g00750, VIT_11s0016g00470 and VIT_17s0053g00700) were identified in this study, among them only the least expressive transcript (VIT_05s0077g01930) was found differentially expressed within T1 infected berries at S1. At most, all these SuSy showed are slightly and non-significantly upregulated at S1 and downregulated at S2 (Additional file 7: Table S6).
Instead, cell wall invertase transcript (VvcwINV: VIT_09s0002g02320), actively expressed at véraison and during late ripening [234] and indicating an important role in the sugar accumulation process, was found to be significantly downregulated by both treatments but only at S1 (Fig. 6; Additional file 1: Table S1).
Six hexose transporters related to glucose and fructose have been identified in grape berries and named VvHT1-6. VvHT1, VvHT2 and especially VvHT3 are generally greatly expressed compared with the other VvHT genes, at all stages of berry development [235]. In the present study and contradictory to previous results reported by Vega et al. (2011) [19], VvHT1 (VIT_00s0181g00010), the first transporter of this family identified in grape [236], and functionally characterized [237] was significantly downregulated by T1 only at S1 (Additional file 1: Table S1). Besides, its transcription factor VvMSA (VIT_18s0072g00380) was non-significantly impacted by GLD treatments (Fig. 6; Additional file 7: Table S6). VvHT2 (VIT_18s0001g05570), associated with véraison and was weakly expressed in ripening berries [34, 235], was non-significantly repressed mainly by T1 at S1. Besides, VvHT3 (VIT_11s0149g00050), also named VvHT7 and which has reduced expression at véraison but high one at both green and ripening stages [234], was slightly and non-significantly upregulated by T2 at both stages and downregulated by T1 at S2. VvHT4 (VIT_16s0013g01950) whose transport activity is restricted to glucose [234], was shown as well non-significantly down- and upregulated at both stages, respectively, by T1 and T2. VvHT5 (VIT_05s0020g03140), was differentially expressed being induced by T1 and repressed by T2 at S1. VvHT5 shows an opposite expression pattern at S2. Lastly, VvHT6 (also named VvTMT1), with an expression activity associated with véraison and during maturation, was described as a tonoplast monosaccharide transporters (TMT) as it has a high similarity with AtTMT2 and serving to hexoses to reach their location storage [235]. A non-significant down modulation was observed for VvHT6/VvTMT1 (VIT_18s0122g00850) by both treatments at S2 (Fig. 6; Additional file 7: Table S6).
Similarly, another VvTMT2 (VIT_03s0038g03940) was non-significantly impacted by GLD. Otherwise, two vacuolar invertases (vINV, VvGINs) VvGIN1 (VIT_16s0022g00670) and VvGIN2 (VIT_02s0154g00090) were differentially expressed within T2 infected berries being induced by T2 but only at S1 (Fig. 6; Additional file 1: Table S1).
Previously, Lecourieux et al. (2014) showed that VvSK1, a protein kinase with sugar signaling function during berry development, positively affects sugar accumulation in grape cells and controls glucose transport through the regulation of four genes that encode the hexose transporters VvHT3, VvHT4, VvHT5, and VvHT6 [238]. Besides, Hayes et al. (2007) suggested that VvHT4 and VvHT5 are not significant contributors to sugar import during berry development giving that these both genes are faintly expressed in Cabernet Sauvignon berry flesh [234]. While, VvHT3 and VvHT6, are considered as the main hexose players of sugar transport and accumulation during berry ripening [234, 238, 239]. Here, VvSK1 (VIT_15s0048g02010) was non-significantly modulated by GLD (Fig. 6; Additional file 7: Table S6). A part from VvHT1 and VvHT5, none of the above transporter hexoses genes was found significantly modulated by GLD. VvHT5 transcripts were reported more abundant at late ripening, but their amount remains low [235] and thus may not contribute to sugar import during berry development specifically at these studied stages S1 and S2. As well, it was determined that both VvHT1 transcripts and protein levels were abundant during the early stages of berry development (pre-véraison) but absent from ripening berries, indicating that it is not directly responsible for the post-véraison sugar accumulation [237, 240, 241]. Moreover, Chen et al. [242, 243] identified a new class of sugar transporters, named SWEETs in plants, animals, and bacteria. In grape, SWEET genes were also identified and characterized, among which some members namely VvSWEET4, VvSWEET7, VvSWEET10, VvSWEET11, VvSWEET15 and VvSWEET17d were shown strongly expressed in berries, thereby suggesting that VvSWEET fulfils important functions in grape berries [244]. 11 SWEET transporters were identified here, among them 4 were differentially modulated and named VvSWEET11 (VIT_07s0104g01340), VvSWEET17d (VIT_14s0060g01910), VvSWEET4 (VIT_14s0066g01420) and VvSWEET12 (VIT_17s0000g00820) (Additional file 1: Table S1). VvSWEET11 was positively regulated by T1 at both stages. Noticeably, VvSWEET4 was highly induced by T1 at S1 and further downregulated by the same treatment at S2 but with a much lower impact. A previous study revealed this glucose transporter localized in the plasma membrane as highly involved in grapevine interaction with Botrytis cinerea being upregulated by inducers of reactive oxygen species and virulence factors from necrotizing pathogens [244]. VvSWEET12 was induced by T2 at S1. Instead, VvSWEET17d was repressed by T1 but only at S1. A recent study characterizing VvSWEET10, as a plasma membrane-localized transporter with hexose affinity and strongly expressed at véraison, prove that this transporter is contributing to glucose and fructose contents and influencing the soluble sugar composition in grapes [245]. Here in, VvSWEET10 (VIT_17s0000g00830) was non-significantly impaired by GLD treatments (Additional file 7: Table S6).
As opposed to Vega et al. (2011) [19], these results indicate the absence of an active and direct effect on sugar transport and metabolism, which confirms also the non-impairment of structural and regulatory genes of the anthocyanin biosynthesis pathway, since a tight positive correlation has been reported between sugars and anthocyanin concentration in berries [246].
{kind=link}