The rapid development of technologies, such as high-throughput sequencing technologies, has provided a huge amount of scientific data, especially in the field of molecular ecology [25]. Moreover, dealing with a huge amount of data as well as using these data to understand functional processes at the community level presents significant challenges. Moreover, the network interactions play an important role in the ecosystem processes and functions. Therefore, based on high-throughput sequencing data, we constructed several networks, investigated the different interactions in the microbial communities of the semi-humid and semi-arid mining ecosystems, and identified the key populations. We also examined the relationships between network structures and soil properties.
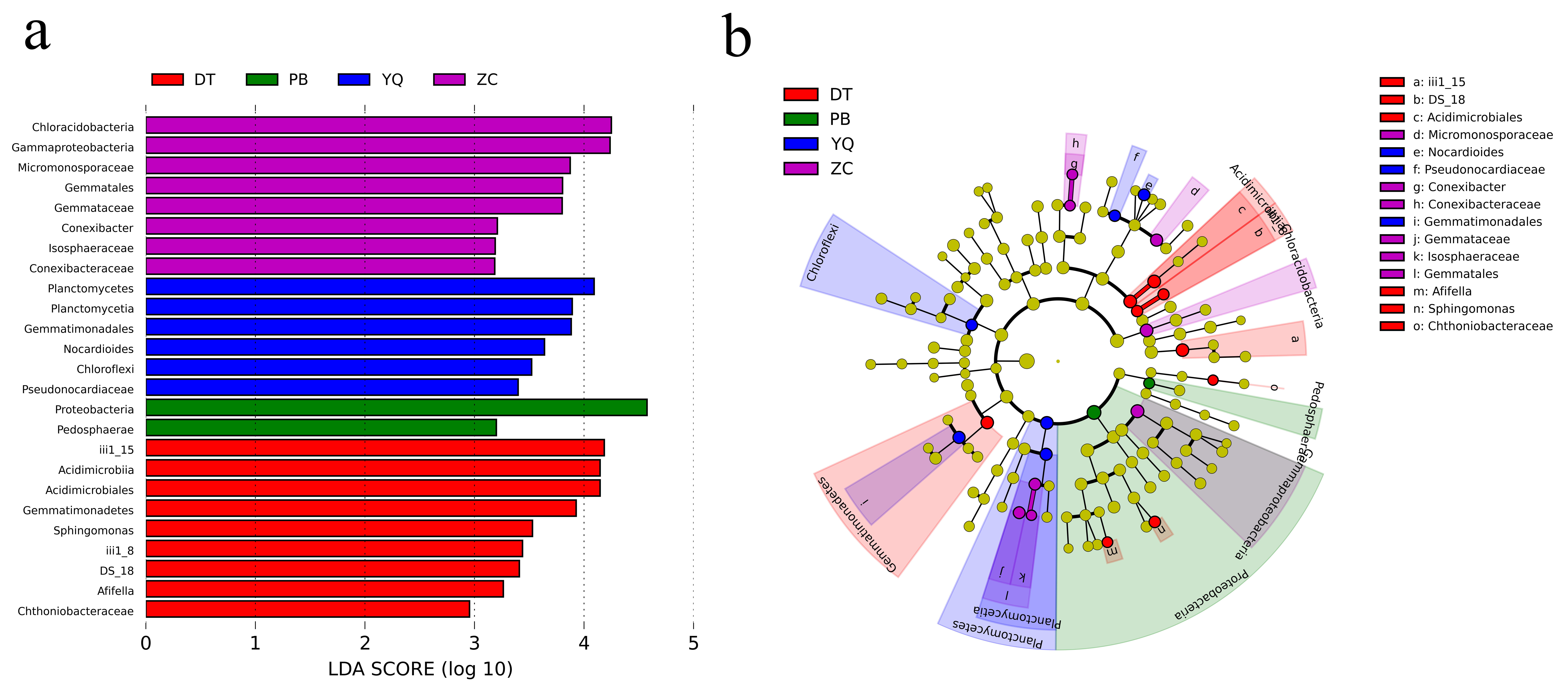
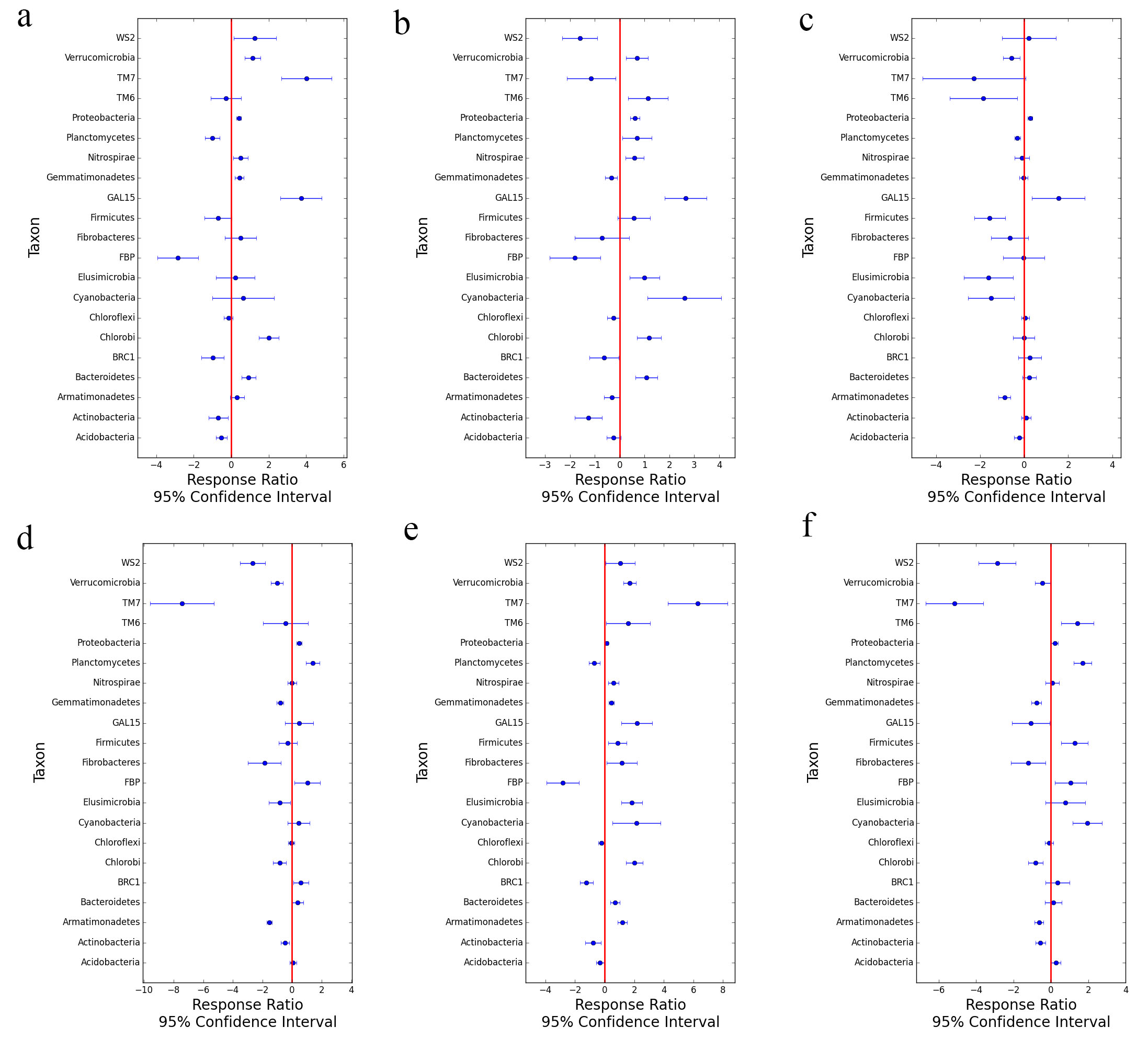
The results of taxonomic composition of microbial consortia in different mining areas (Table 1, Fig. 1) suggested that, the observed species changed across long distances, which might imply that there were some new species generated with changed locations. Although the disturbed mining soil environment might pose a challenge for some species in the soil, it still stimulated new microbial species, especially for bacteria that can adapt to special reclaimed environment. From the semi-humid locations to semi-arid areas, soil microbial diversity showed the decreasing trends, which might suggest that some special environment might be formed in the regional mining damaged sites, and caused some bacteria to die. The most soil microbial phyla represented in this study belonged to 13 major phyla. Nevertheless, the species distributions on the phylum level were different for the four mining areas. Regarding the temporal variation in Fig. 2, the PCA and NMDS results indicated that the microbial communities changed throughout the spatial distance. Figure 2 displayed that the PB group was far away from the DT group, while the distance was near between ZC and YQ. Although the percentage of PC2 (principal component) explanation was just 11%, the result obtained on this axis still might be reliable in interpretation (Fig. 2a). This result in Fig. 2b might indicate that the microbial structure become more different as the changing location from semi-humid to semi-arid mining areas, which was in line with the research investigated by Helingerová et al [17]. Moreover, methods of response ratio calculation (RRC), and linear discriminant analysis Effect Size (LEfSe) were also used to analyze the differences among the soil community structures of the four mining areas (Fig. S5-S6 in Additional file 15–16).The RRC and LEfSe results also implied the gaps among the four groups, which confirmed that the observed changes of microbial community were significantly impacted by the changing spatial locations. Moreover, in spite of the soil microbe have been significantly disturbed or destroyed by mining activities, especially in the semi-arid areas, the soil microorganism might resuscitate or restore by the interaction by themselves.
In this study, we also analyzed the microbial interactions in different mining areas using the method of molecular ecological network analysis. The network properties changed across all four mining areas (Table 2) from semi-humid to semi-arid areas, and the species involved in the microbial interactions also changed, as demonstrated by variations in dominant phyla (high node degree) (Fig. 3–5). In the networks, community stability was higher with increasing complexity. The simple network structure (no connectors or module hubs and more sparsely distributed species) and low competitive connections may have caused a negative effect on biogeochemical functions, indicating an unstable and vulnerable microbial community when other disturbances occurred. Microorganisms under this condition might have been specialized to the local environments and thus sensitive to environmental changes. Furthermore, the results (Table 2 and Fig. 3) indicated that the network interactions for some microbial groups were more complicated in the PB network located in the semi-humid area, although the microbial community diversity in this network was the poorest. The nature parameters in PB and ZC are totally different from those of YQ and DT. Moreover, this result implied that different natural condition might have significantly affected the microbial community structure and their network interactions in different ways.
The networks obtained showed the general features of many cellular networks, such as modular, small world, or scale free [26]. A small-world pattern contributed to the efficient communication among different members in a community and could quickly respond to external environmental changes, such as mine subsidence, subsidence cracks, landslides, or soil reclamation. Closeness centrality is based on the average shortest paths and thus reflects the central importance of a node in disseminating information. Complex networks with greater connectivity are more robust to environmental perturbations than simple networks with lower connectivity [27, 28]. In this sense, the higher complexity of the PB and ZC networks suggested that (Table 2), as different taxa were complementary, the microbiome in the eastern semi-humid mining areas with a high groundwater level was more resilient to environmental stresses, such as mine subside or land reclamation activities. The result might imply that the network structural complexity might be related with geographic location and environment. Further studies are necessary to corroborate this observation.
Moreover, we considered the OTUs with the highest degree and highest closeness centrality, and the lowest betweenness centrality scores to be the keystone taxa [29]. Keystone taxa are highly connected taxa that play important roles in the microbiome, and their removal can cause significant changes in microbial composition and functioning [30]. Although previous studies have reported keystone taxa in various environments, reports on keystone taxa in the disturbed mining areas have been limited [31–34]. As found in this study, key populations can be distinguished according to their network profiles and module memberships. Networks in the semi-humid mining areas with a high groundwater level, such as PB and ZC, showed that the keystone taxa belonged to the microbial phyla Acidobacteria, Gemmatimonadetes, Chloroflexi, and Proteobacteria, whereas Actinobacteria, Acidobacteria, and Chloroflexi were the key species in the YQ and DT networks from semi-arid mining areas (Fig. 2–5; File S1-S4 in Additional file 1–4). Although the YQ mining area is far away from the PB and ZC mining areas, the most abundant phylum was the same (i.e., Proteobacteria), whereas Actinobacteria was most abundant in the DT mining area, despite the fact that the two mining areas are closer to each other (Fig. 8). This result might indicate that no direct relationship existed between the location sites and microbial abundance. As we know that Proteobacteria is distributed widely around the world. It has an aerobic bacterium that is capable of degrading a variety contaminants as well as some bacteria producing several oxidases that oxidize diverse compounds [35]. Proteobacteria has a highly diverse physiology and is distributed in almost all of the different ecological environments. Mining areas are complicated and contain surface subsidence, cracks, landslides, reclamation, and restoration areas. This complicated condition might result in a suitable environment for Proteobacteria, which may have made Proteobacteria the dominant bacteria. Actinobacteria are ubiquitous gram-positive bacteria, and have a characteristic filamentous morphology, which might be the reason for its high abundance in the semi-arid DT mining area. In addition, Actinobacteria have a variety of important functions that make them useful and powerful in soil and marine environments, including degradation of organic substances. In the adverse and comprehensive semi-arid mining areas, the existence of Actinobacteria being the dominant microbe might help improve the soil quality. Acidobacteria play a significant role in soil ecological processes, and this diverse phylum is distributed widely throughout various natural environments [36, 37].
On the other side, the abundances of key taxa Chloroflexi and Gemmatimonadetes were low, which suggested the lack of a direct relation between abundance and key functional importance. Chen et al. (2017) have reported that Chloroflexi increased when the environment became more anaerobic. It is possible that mining areas have many kinds of environments, such as surface subsidence or cracked areas, which are suitable for the Chloroflexi [38]. Even though the ecological function of Chloroflexi was not clear, this phylum was still the keystone microorganism in the four mining areas. Recently, phylum Gemmatimonadetes has been described as a bacterial group whose members are widespread in soil habitats. Its cultured representative genus is Gemmatimonas aurantiaca, which has been isolated and able to grow under not only anaerobic conditions but also aerobic conditions [39, 40]. This finding might suggest that Gemmatimonadetes could be a suitable phylum in complicated mining areas that contain aerobic and anaerobic environmental habitats. This might be the reason why the Gemmatimonadetes was the keystone taxa. Tobin-Janzen et al. [41] reported that Nitrospira was the dominant genus of bacteria in soil samples form an underground coal-mining fire (Pennsylvania, USA). Sun et al. [42] reached a similar conclusion that Nitrospira accounted for the highest proportion in the soil samples from China. However, what these past studies found was different from the results from our study. Ezeokoli et al. [43] have investigated the microbial community in opencast coal mines but did not study the keystone taxa. Their results showed that microbial communities in mining areas have been impaired and have had negative effects on soil biological processes, especially nutrient cycling and ecosystem sustainability.
In this study, the connection between two OTUs indicated that the two OTUs might respond to a common environmental parameter. Then characterization of the OTU connections in modules could be used to describe these interactions among the microbial communities [44]. Also, it might be suggested that the same underlying factors motivated changes in OTU abundances with strong module memberships. Therefore, OTUs with strong module memberships should have some physical or functional relationships in the community. As shown in this study, module memberships, topological roles, and phylogenetic relationships have provided some information to identify the key OTUs. Thus, the interactions and ecological roles of these microbial communities in mining areas might provide insight for mining activities in China, especially for ecologically fragile and vulnerable areas. For the first time, this study presented different network interactions among soil microbial communities in semi-humid mining areas with high groundwater levels and semi-arid mines.
In fragile ecological systems, understanding how the soil microbial communities respond to external environmental changes, in particular, for anthropogenic change, is significantly important [45]. In this study, the method of network analysis revealed an appropriate way to discover how environmental changes affected microbial communities. Previous studies have shown that when the external environment changed, such as variations in soil properties, the diversity of microbial communities changed, which may be correlated with disturbances in soil characteristics [46–48]. Additionally, soil factors, such as pH, moisture content, total carbon content, and organic matter, reportedly have had a greater impact on soil bacterial community structure and diversity in the ecological restoration of mining areas. For example, Xiao et al. [49] have reported that soil microbial activity was affected by soil factors to different degrees, and that soil microbes played a critical role in the recycling of soil nutrients and soil fertility. Pille Da Silva et al. [50] found that soil microbiological attributes affected microbial biomass carbon and microbial basal respiration. The microorganism could increase soil quality and restore biological diversity in the coal-mining area. Understanding this relationship between a microbial community and soil properties is critical to the ecological restoration of coal-mining areas. Bi et al. [51, 52] found that the arbuscular mycorrhizal (AM) fungal community was influenced by mine slope position and subsidence. Their study clarified that AM fungal ecological function could potentially help in vegetation restoration and reduced erosion in coal-mining areas.
In our study, we identified strongly significant or significant correlations between the node connectivity in the module and the selected environmental variables, such as AAT, soil pH, SOM, and ammonium nitrate content, for the PB network. Fernández-Montiel et al. [53] showed that soil pH could change the slightly acidic environment to an acidic condition. Results from 12 sites following mining activities of different lengths of time of reclamation suggested that soil microbial abundance, taxonomic diversity, and functional diversity could be improved by increasing the number of reclamation years [54]. A redundancy analysis revealed that soil pH was significantly important in microbial metabolic structure and bacterial genetic assemblages. This finding was similar to our results that soil pH value was significantly correlated to different species of module 6 just in PB network (Fig. 6), especially for the OTU_19170 (Koribacteraceae), which belonged to Acidobacteria (File S1 and Fig. S1 in Additional file 1 and 5). Sáenz de Miera et al. [55] presented the finding that subgroups of Acidobacteria showed a significantly positive relationship with soil pH value. Soil organic matter is always represented as an important indicator to estimate the soil carbon storage and to evaluate soil quality. Disturbances introduced by mining activities might affect the activity of soil microbes, thus affecting the SOM content. The results in this study also showed that the SOM had a significant relationship with module 6, of which the important nodes OTU_4611 (Burkholderiales) and OTU_8175 (Burkholderiales) belonged to phylum Proteobacteria, which was the keystone phylum in the PB network (File S1 and Fig. S1 in Additional file 1 and 5). Therefore, all of these results may have suggested that pH value and SOM revealed a complicated relationship with soil microbial communities, in particular with the keystone species in PB network. In a sense, these results confirmed that the method of network analysis was effective and feasible to analyze the relationship between environmental factors and microbial community structures.
The AAT and EC value was significantly related to two modules in the ZC network. During the succession of land following coal mining, aggregate stability and organic matter increased, whereas EC value decreased. Other researchers have examined the soil bacterial characteristics of 21 coal-mining sites [56]. One result was that the bacterial species composition was significantly correlated with the soil EC value, which was similar to conditions in the ZC network. Our results showed that the soil EC value was significantly correlated with different species of module 1 (Fig. 6), especially for the OTU_34138 and OTU_8126 (Acidobacteria), which belonged to keystone species in the ZC network (File S2 and Fig. S2 in Additional file 2 and 6). The EC value showed as a kind of soil-leaching solution reflected the water-soluble salt content in soil. Once the soil was disturbed by mining activities, the solubility of calcium carbonate or magnesium carbonate in the soil might have been affected, and then the water-soluble salt content in the soil-leaching solution changed, which influenced the microbial communities. Sun et al. [42] have found that the distribution of bacteria was affected primarily by SOM, AK, and AP in similar coal-mining areas. The location map in Fig. 8 shows that PB and ZC are close to each other, and belong to the semi-humid area. Moreover, both of them are locating in the coal-mining areas with high groundwater levels. For these areas, the soil was affected by a secondary anti-alkali and heavy metal migration problem. In the future, we need to investigate additional properties, such as heavy metal contents.
In the DT network, the soil variable pH and AP showed a significant correlation with the module, whereas we did not identify a significant correlation between the modules and soil variables in the YQ network. This result implied that pH and AP value might have played an important role in the DT network structure. In the DT network, pH showed a positive relationship with the phyla Acidobacteria. Notably, the important nodes in module 7 all belonged to Acidobacteria (File S4 and Fig. S8 in Additional file 4 and 8). This suggested that Acidobacteria was significantly correlated with the soil pH value. Ma et al. [35] have reported similar results that the abundance of Acidobacteria changed with variation in the soil pH value. In this study, this result indicated that external environmental variables affected the network interactions among different microbial groups and that such changes may be related to soil properties, such as pH value. These results also indicated that both pH value and mine activities affected the microbial and network structures.
Furthermore, AP showed a significant correlation with module 12 in the DT network, and the important nodes in module 12 (Fig. 6) belonged to Gemmatimonadetes (OTU_34734, OTU_13954, and OTU_19203) and Chloroflexi (OTU_129) (File S4 and Fig. S8 in Additional file 4 and 8). This indicated that Gemmatimonadetes and Chloroflexi were significantly correlated with soil AP value and that Chloroflexi was the keystone species in the DT network. Furthermore, the presence of microbes in the same module implied that these microbial populations compartmentalized with each other to survive in response to disturbances caused by mining activities. It is well know that phosphorus is one of the most indispensable nutrient elements for soil development. It can be easily fixed in the soil, although its utilization rate is low. Moreover, phosphorus is a necessary element for microbial metabolism—for example, some soil microorganisms may produce acidic substances through metabolism, and then dissolve some insoluble phosphates and apply them to their own metabolic processes. All of these results could indicate that the soil phosphorus content might be correlated with keystone species. We speculated that the disturbed environmental factors influenced the microbial composition, thus influencing the AP content. In this study, we did not find any significant correlation between environmental factors and network modules in the YQ network, which implied that we need to examine and include additional environmental factors in this analysis, or developed a new method to prove this relationship.
Combined the results of mantel test, correlation test, CCA and RDA together, we found that, in spite that the environmental variables such as AP, AK and EC showed significant effects on the microbial communities, the explanation percentages in VPA plot (Fig. 7) were very low. However, the natural factor AAT could explain 13.725% on the effect, which might suggested that natural geographic condition influence the microbial community structures. On the other side, soil pH value and SOM are well known being the key environmental factors which affected the soil bacterial communities [46, 53–55]. However, on the phylum level, pH and SOM showed no effects on the microbial community structures, which might imply that natural geographic factors, such as the spatial distance (from semi-humid to semi-arid locations) provide a key role in soil microbial compositions.
Information on the common presence of bacteria related to keystone microbes, however, is still insufficient for these networks. We still could not identify the exact keystone species and their differentiation between the semi-humid and semi-arid mining areas, which may have mitigated the effects of soil disturbance and accelerated the restoration of mined soil. Moreover, the high-throughput 16S RNA gene sequencing provided extensive information about only the taxa present in bacterial communities in disturbed mining areas, but did not provide enough insights about the functional roles of these keystones, which is essential for ecological restoration. The ecological function network analysis and more extensive research of metabolism should be investigated in the near future using the Geochip technology.
{kind=link}
{kind=link}