Effects of calcium on the karst adaptive characteristics of Camellia
Abiotic stress signals can cause fluctuations of cytoplasmic calcium ions and changes in phosphorylation forms of some proteins. Calcium ions mediated by calcium channels, change the concentration of calcium ions in cytoplasm and amplify the stress signals with intracellular calcium signal receptors [22]. However, high calcium stress can destroy the cytoskeleton and biofilm structure, resulting in intracellular metabolic imbalance.
In order to study the adaptability of the karst environment with high calcium content of Camellia limonia, we first processed Camellia limonia with high calcium content and take Camellia nitidissima as the control, the results showed that the adaptability of Camellia limonia to the environment with high calcium content was stronger.
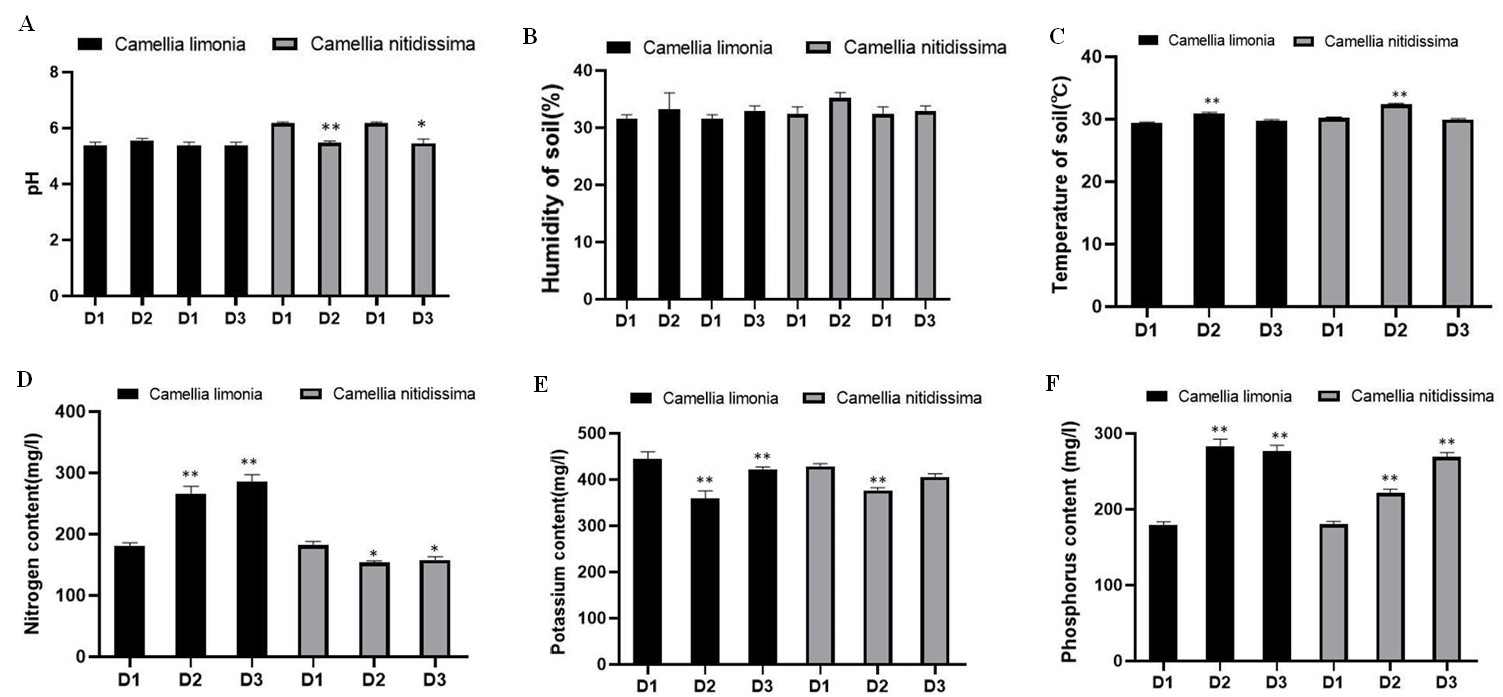
Under the calcium stress, the phenotypes of Camellia nitidissima and Camellia limonia became different. The leaves of Camellia nitidissima turned yellow and wilting, while the leaves of Camellia limonia kept relatively healthy (Figure 1). The SEM and microscopic observation also showed that the leaves and roots of Camellia limonia are less wrinkled and dehydrated than Camellia nitidissima (Figure 2). The stomatal conductance and transpiration rate of the two kinds of Camellia showed a downward trend. The stomatal conductance of Camellia nitidissima decreased more than the one of Camellia limonia. While the reduce degree of transpiration rate Camellia limonia is less than in Camellia nitidissima. The chlorophyll content of Camellia limonia increased by 53.12% and Camellia nitidissima decreased by 56.91%. The net photosynthetic rate of Camellia limonia was more stable than Camellia nitidissima. The average net photosynthetic rate of Camellia limonia and Camellia nitidissima was 3.0853 μmol CO2 m-2s-1 and 2.4456 μmol CO2 m-2s-1, respectively. Stomatal conductance affects the oxygen and carbon dioxide, indicating that the water might be the more important factor influencing the net photosynthetic rate when gas were equivalently affected [23]. According to the data, the average leaf humidity of Camellia limonia is 21.29% RH, Camellia nitidissima leaf humidity averages of 17.88% RH. The average transpiration rate and leaf temperature of Camellia nitidissima were 0.64 mmolH2Om-2s-1 and 31.94 ℃,Camellia nitidissima were 0.92 mmolH2Om-2s-1 and 30.95 ℃, suggesting the water content of Camellia limonia is more than Camellia nitidissima (Figure 3). According to the results above, Camellia nitidissima has relatively stronger adaptability to the environment with high calcium.
The soil moisture and temperature of the two kinds of Camellia did not obviously change. The pH value of the soil of Camellia limonia decreased slightly. In addition, the nitrogen and phosphorus content in the soil of Camellia limonia increased by 58.60% and 53.99%, respectively, while the nitrogen and phosphorus content in the soil of Camellia nitidissima decreased by 13.01% and increased by 49.63%, respectively (Figure S1).
Available nitrogen and phosphorus are the main nutrient elements that restrict plant growth, while plant growth is mainly restricted by nitrogen elements under low ratio of nitrogen and phosphorus [24]. Under high calcium stress, nitrogen content of Camellia limonia leaf increased by 14.44% and the available nitrogen in soil of Camellia limonia is increased by 58.60%, while nitrogen content of Camellia nitidissima leaf decreased by 28.57% and the available nitrogen in soil of Camellia nitidissima decreased by 13.01%, suggesting Camellia limonia has better nitrogen use efficiency (Figure 3), which may also help Camellia limonia survived in high calcium environment.
Metabonomics of Camellia leaf in karst high calcium environment
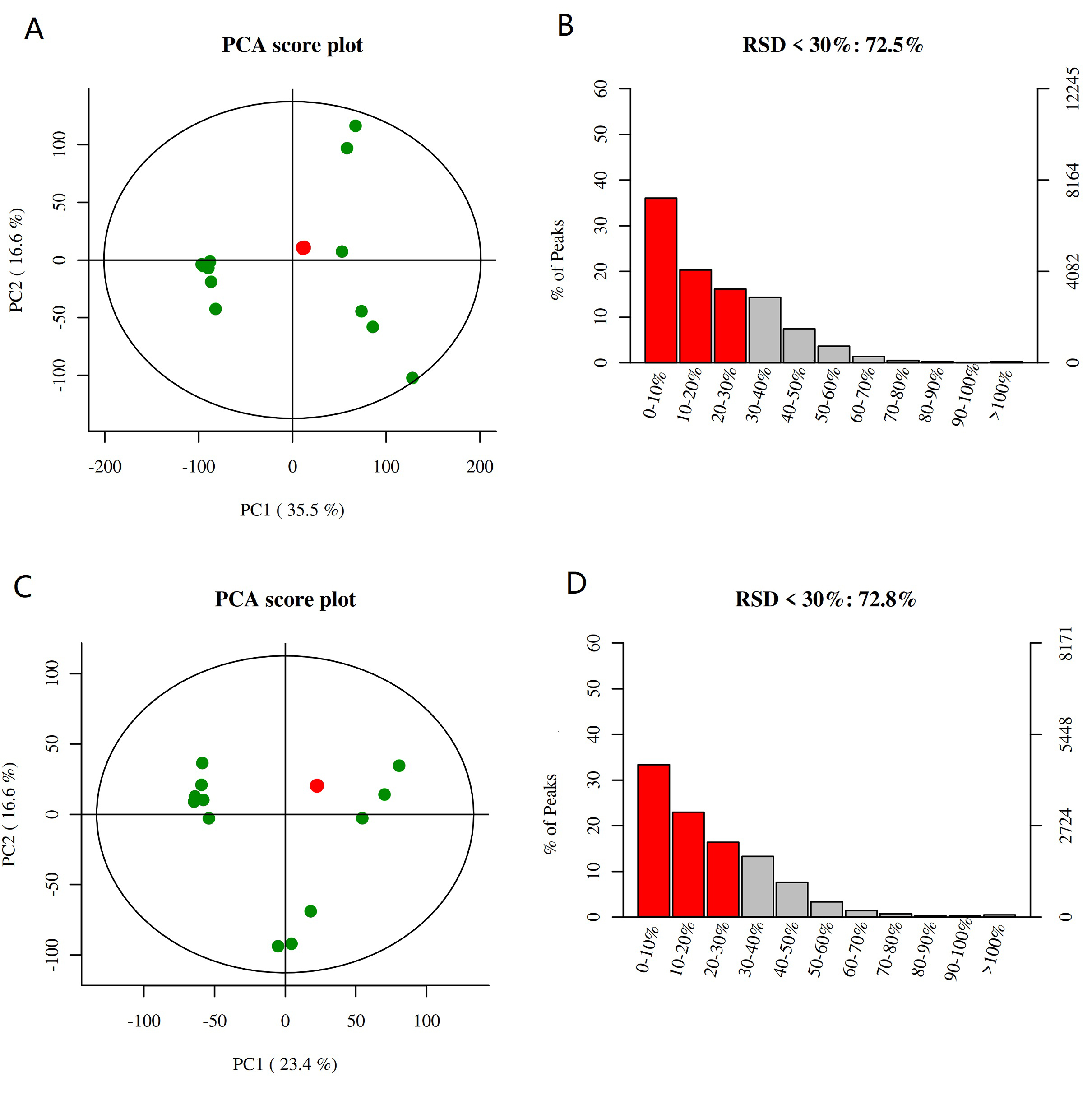
In order to analysis the different metabolites of Camellia leaf in karst high calcium environment, the metabonomics were conducted. The PCA score chart showed the degree of aggregation and dispersion of Camellia limonia and Camellia nitidissima (Figure S2). The distribution points of Camellia limonia are relatively close, indicating that the composition and concentration of the variables/molecules contained in Camellia limonia are closer.
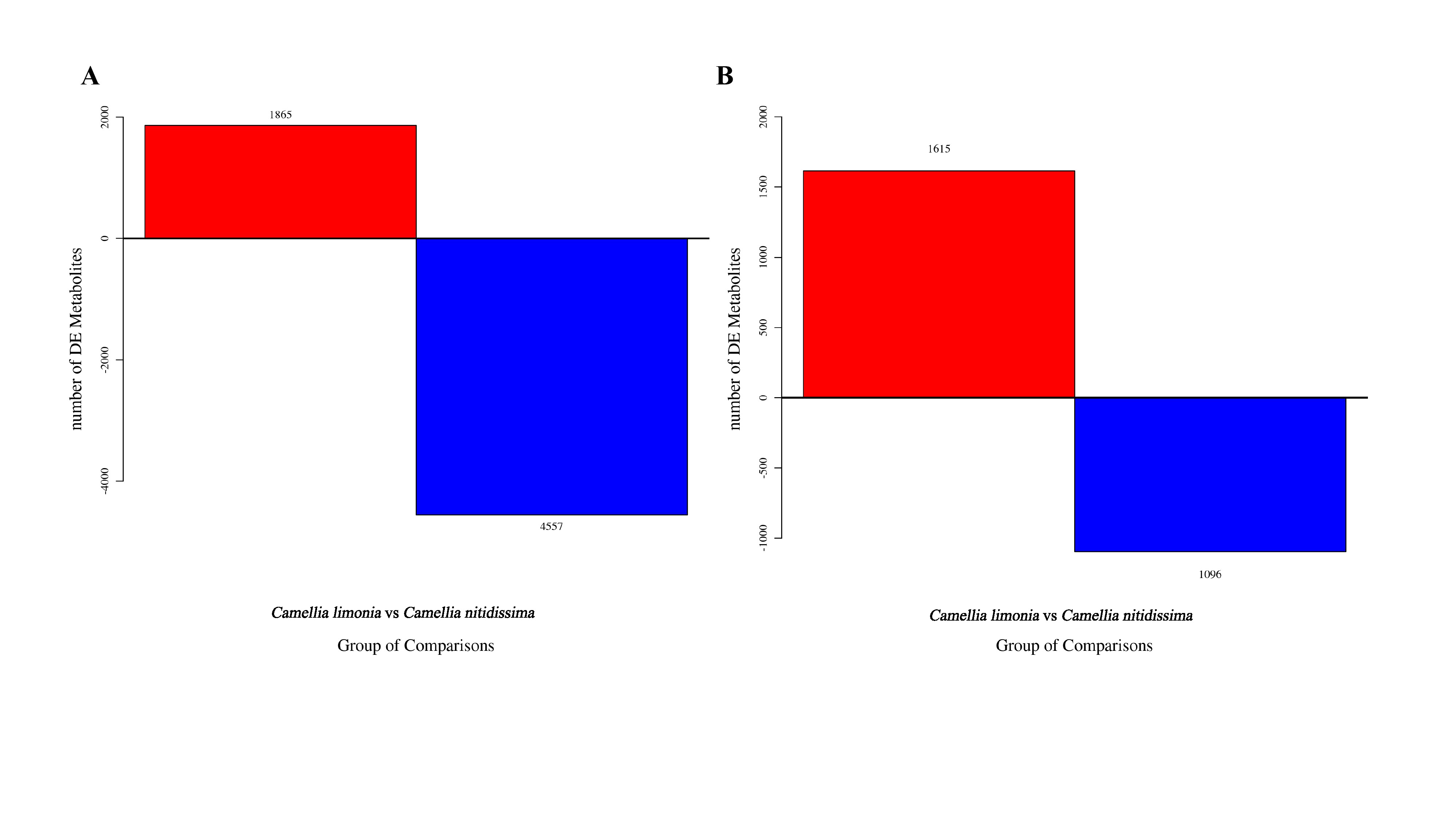
A total of 9133 metabolites were identified through mathematical statistical analysis and screening, of which 6422 were in the positive ion mode and 2711 were in the negative ion mode (Figure S3). 8966 differential metabolites discemed include metabolites of Camellia limonia and Camellia nitidissima. The accurate molecular weight of the metabolites (molecular weight error <15ppm) were confirmed and the fragment information were divided according to the MS/MS mode. We identified 220 different metabolites in HMDB (Human Metabolome Database, HMDB), metlin, massbank, lipiduaps, mzdound databases and obtained the statistical analysis results (Table S1). Under the high calcium stress, Camellia limonia has 53 metabolites up-regulated and 167 metabolites down-regulated, of which the main up-regulated metabolite is kaempferol-3-o-rutinoside and tyrosol. Camellia nitidissima has 167 up-regulated metabolites and 53 down-regulated metabolites, of which the main metabolites up-regulated are n-acetylhistidine, myristic acid, and pelargonin (Figure 4). The five different metabolites mentioned above have the largest significant difference. The content of these metabolites were changed to affect the metabolic process of Camellia, indicating that the significantly different metabolites mainly affect the growth of Camellia limonia and Camellia nitidissima after the treatment of high calcium stress.
Among them, kaempferol-3-o-rutinoside, tyrosol, 6-o-methyldeacetylisoipecoside,n-acetylhistidine, myristic acid and pelargonin are positively correlated. But the three metabolites of Camellia limonia were negatively correlated with the three metabolites of Camellia nitidissima (Figure 5). It is inferred that the up-regulated metabolites of Camellia limonia had the opposite trend to the up-regulated metabolites of Camellia nitidissima, indicating that Camellia limonia have different metabolite trends in high calcium stress.
MetPA is a part of metaboanalyst (www.metaboanalyst.ca) and is mainly based on the KEGG metabolic pathway. The MetPA related metabolic and KEGG pathway analysis show 42 main metabolic pathways. The most important is biotin metabolism, tyrosine metabolism, flavone and flavonol biosynthesis, inositol phosphate metabolism, alpha-linolenic acid metabolism, ubiquinone and other terpenoid-quinone biosynthesis, phenylpropanoid biosynthesis, lysine biosynthesis, phenylalanine metabolism, glyoxylate and dicarboxylate metabolism, pantothenate and CoA biosynthesis, arginine and proline metabolism and other metabolic (Table S2). The pathways with most significant differences are tyrosine metabolism, flavone and flavonol biosynthesis, phenylpropanoid biosynthesis and biotin metabolism. And more differential metabolites are enriched in these four pathways, indicating that these pathways are more sensitive to high calcium stress for Camellia limonia.
Metagenomics of soil in karst high calcium environment
Under biotic and abiotic stresses, plants can actively change their own physiological conditions and exudates of their roots. That also has effects on the related microbial communities. Consequently, this directly or indirectly affects the health of plants. The initial central microorganism and exudates of plant root together form a microbial community, including plant central flora, core flora and other microbial communities. Compared with the plants under high calcium treatment, the root microbial populations of the plants in normal condition are different. Therefore, these different populations could affect the growth of plants. Metagenomics analysis of the soil of Camellia limonia and Camellia nitidissima root were conducted to study the effect of microbial communities under high calcium stress. The result showed that tyrosine metabolism, galactose metabolism, inositol phosphate metabolism, biotin metabolism, phenylpropanoid biosynthesis, arginine and proline metabolism were significantly affected. The microorganism regulated these metabolic pathways. At the same time, they regulate the expression of responsive proteins through responsive genes. Finally, these microorganisms influnce the metabolic pathways of Camellia.
The software Velvet was used to evaluate the assembly results, including the statistical table of assembly results, the distribution chart of the cumulative length of contigs and the distribution map of GC content (Table 1).

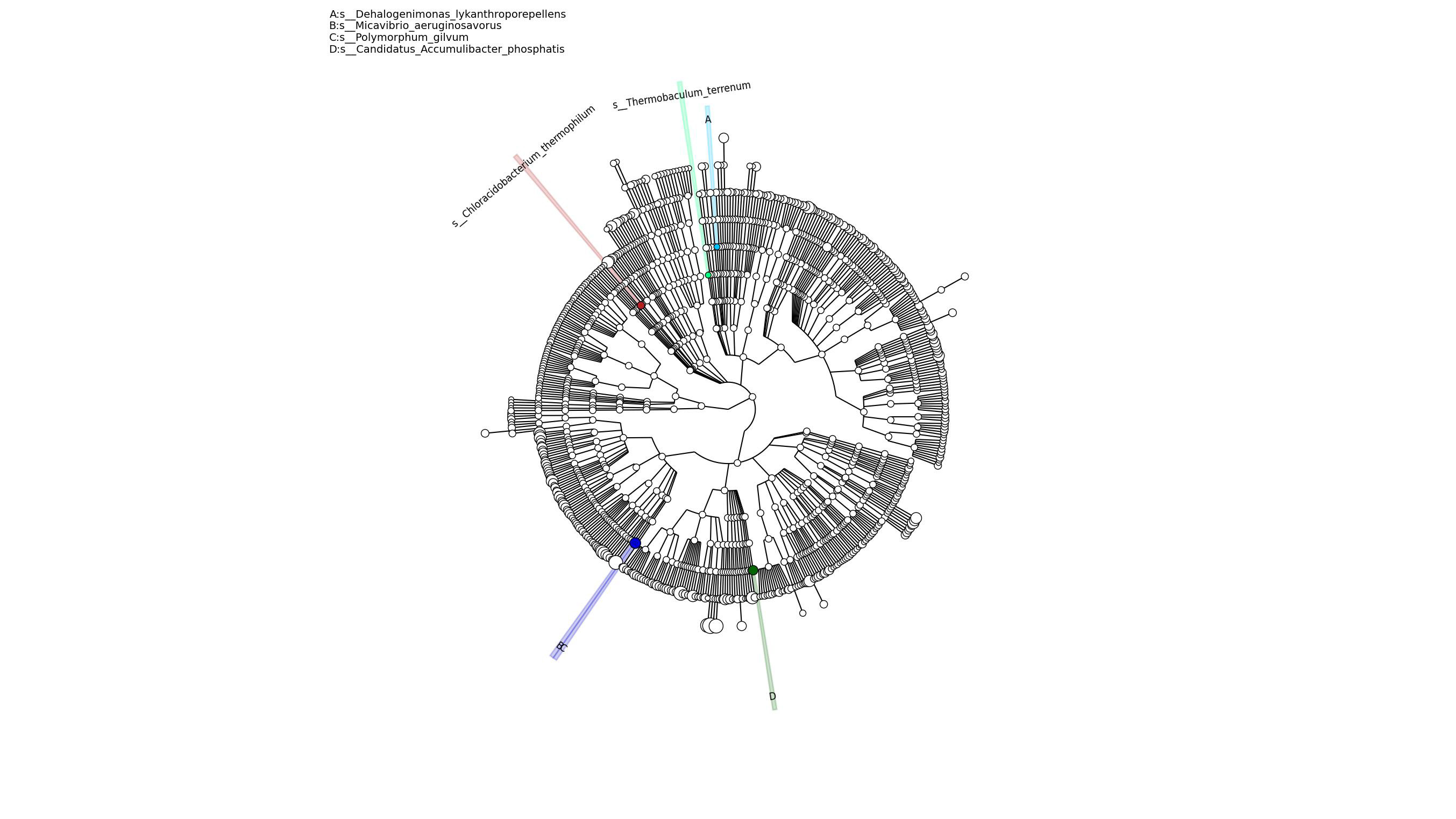
We examined nearly 20,000 genes from each sample of rhizosphere microbes. And we found that reduced pathogenicity and loss of toxicity were the most important factors. In order to get a more detailed classification result of species, we match the sequence with the bacterial reference sequence or the virus reference sequence. Moreover, the MetaPhlAn was used to comment the species classification (Table 2). In all samples, rubrivivax is the main member. In root soil of Camellia limonia, rhodanobacter, intrasporangium, microbacterium, ilumatobacter are taking disadvantages. Different from Camellia limonia, the Candidatus_Solibacter was the dominant species of Camellia nitidissima (Figure 6). Further, in order to reveal the distribution and evolution relationship of species, phylogenetic analysis was conducted (Figure S4). The three colors represent the most important four species (dehalogenimonas lykanthroporepellens, micavibrio aeruginosavorus, polymorphum gilvum and candidatus accumulibacter phosphatis) and their evolution relationship. The above results about the metagenomic analysis indicated that, compared with Camellia nitidissima, the types of soil microorganisms of Camellia limonia under high calcium stress were different. And these microorganisms are related to the salt tolerance and drought resistance of plants.

Camellia limonia adaptive regulation to karst
In order to study the karst adaptation of Camellia limonia, we conducted the integrative analysis of metabolomics and metagenomics. The result showed that flavone and flavonol biosynthesis pathway which is one of the most affected pathways can be regulated by apigenin, kaempferol, astragalin, isoquercitrin metabolites and TT7, UGT78D1, UGT78D2 genes. This metabolic pathway involves the synthesis of flavonoids. Flavonoids have functions in drought and salt resistance [25], so flavonoids may also contribute to the karst environment adaptation for Camellia limonia. Flavonoids include kaempferol, rutin, hesperidin, anthocyanin, etc. Among them, kaempferol has the largest difference in content between Camellia limonia and Camellia nitidissima , and the content of Camellia limonia is greater than Camellia nitidissima. Therefore, Camellia limonia grow better than Camellia nitidissima under the same calcium stress, which indicated that kaempferol may play a critical role in the high calcium tolerance.
The citrate cycle (TCA cycle, Krebs cycle) is an important aerobic pathway for the final steps of the oxidation of carbohydrates and fatty acids. The citrate cycle is also an important pathway for differential metabolites. In the relative pathway of TCA cycle, the sdhA, frdA genes regulate the synthesis of succinate dehydrogenase/fumarate reductase-flavoprotein subunit. Compared with Camellia nitidissima, the sdhA, frdA were differently expression genes in Camellia limonia, which suggested that this two genes might simultaneously regulate the synthesis of fumarate and succinate and further affect the high calcium tolerance of Camellia limonia.
{kind=link}
{kind=link}
{kind=link}
{kind=link}