Malaria affects half of the world’s population and constitutes one of the leading causes of death in developing countries. A parasite, Plasmodium falciparum, is the primary cause of malaria. Artemisinin is a sesquiterpene lactone isolated from the aerial parts of Artemisia annua L. (1, 2). Artemisinin-based combination therapies (ACTs) are the most efficient treatment against malaria (3). Beside the anti-malaria activity, artemisisnin has been proven as an anticancer drug against colon, breast and prostate cancer (4–6), leukemia (7), and hepatitis B (8). So far, the only commercial route for artemisinin supply is plant extraction, but unfortunately, the yield is low and not cost-effective (9). Metabolic engineering and synthetic biology provide an alternative and scalable route for the heterologous bioproduction of interesting natural products in microbial hosts (10).
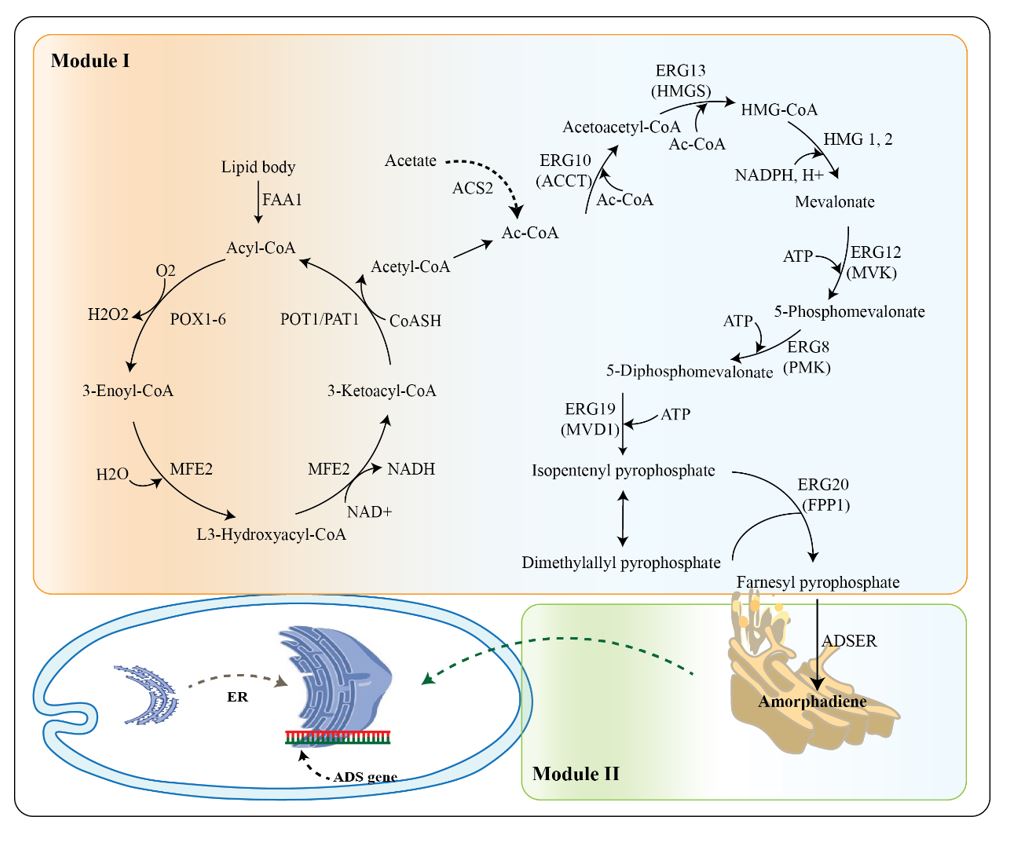
Amorphadiene is an olefin sesquiterpene; the first dedicated step to artemisisnin synthesis in plants is by cyclization of farnesyl-pyrophosphate (FPP) mediated by amorphadiene synthase enzyme (ADS) (11). Isopentenyl pyrophosphate (IPP) and its isomer dimethylallyl pyrophosphate (12), which are biosynthesized from acetyl-CoA through mevalonate pathway, are converted to amorphadiene by the action of the FPP enzyme (9). Geranylgeranyl diphosphate (12) is another intermediate in the mevalonate pathway that, in addition to participating in carotenoid biosynthesis, can also synthesize farnesyl diphosphate from geranyl diphosphate (GPP) and IPP (13). Intending to provide direct and scalable access with cost that is comparable to the agriculture-sourced artemisinin, many researchers sought heterologous production of amorphadiene in various microbial hosts such as Saccharomyces cerevisiae (14, 15), Escherichia coli (9, 16, 17), Bacillus subtilis (18), Azospirillum brasilense (19), and Yarrowia lipolytica (20). However, their attempts could not meet the increasing demand for artemisinin-based combination therapy (ACT) and amorphadiene/artemisinin is not yet commercially available (21).
Yarrowia lipolytica has been considered as a generally regarded as safe (GRAS) platform for the bioproduction of various natural products (22) with its high secretion capacity, strong acetyl-CoA and malonyl-CoA flux (23–26), and a large collection of genetic tools (25–28). Y. lipolytica is a dimorphic, non-pathogenic ascomycetous yeast with a superior host for metabolic and genomic characteristics. It can be a great platform for cost-effective production of biochemicals derived from fatty acids, lipids, and acetyl-CoAs (29, 30). In our previous study, by harnessing the innate mevalonate pathway, we introduced Y. lipolytica as a promising microbial host with a significant production capacity of amorphadiene in a monoculture (20).
In addition to improving the metabolic engineering techniques for bioproduction of a wide variety of value-added biochemicals through engineering a single microorganism, this methodology is still challenging. Increased requirements for fulfilling complicated biosynthetic pathways and reaching efficacy are two critical issues facing monoculture fermentations. They have been considerably circumvented by employing modular co-culture approaches (31, 32). With manipulating the mixture of compatible hosts and nutrients, co-culture strategy provides a platform that absorbs cheaper substrates, improves cell growth without complex treatments, and extends the yield and spectrum of final product/s (33). The optimized Y. lipolytica- Chlorella pyrenoidosa co-culture increased carbon and nitrogen assimilation and drove the carbon flow to a higher yield of microbial biomass such as lipoid, carbohydrates, and protein (34). Consuming lactate as a carbon source and decreasing lactic acid in the media, Y. lipolytica participated as a partner accelerated the Lactococcus lactis growth and accumulation of nisin up to 50% in Y. lipolytica-L. lactis co-culture (35).
To expand the biosynthesis of amorphadiene, a co- culture of two Y. lipolytica strains, Po1g, and Po1f, was investigated in this study. Although Y. lipolytica produces a high amount of acetyl-CoA and malonyl-CoA, our previous attempt demonstrated that harnessing lipogenic acetyl-CoA pathway could improve the bioproduction of amorphadiene (20). So, we first engineered the Po1g strain to improve its secretion capacity. Then to maximize the HMG-CoA uptake and reduce the byproduct production, the amorphadiene biosynthesis was localized in the endoplasmic reticulum (ER) (36). In addition to studying multiple sugar sources, the enlargement of the endoplasmic reticulum was investigated to provide a larger factory to improve the amorphadiene bioproduction. This research will provide more comprehensive insight into the application of Y. lipolytica co-culture to promote cost-effective bioproduction of natural products.
{kind=link}