On 30/01/2020, a 47-year old woman from Wuhan, Hubei province, China presented to an emergency department in Melbourne, Australia. Her symptoms commenced 4 days earlier with lethargy, sore throat, dry cough, pleuritic chest pain, mild dyspnoea and subjective fevers (Fig.1a). She had travelled 11 days prior to presentation from Wuhan via Guangzhou to Australia. She had no contact with the Huanan seafood market or known COVID–19 cases. She was otherwise healthy, non-smoker, taking no medications. Clinical examination revealed a temperature of 38.5C, pulse rate 120 beats/minute, blood pressure 140/80 mmHg, respiratory rate 22 breaths/minute, and oxygen saturation 98%, while breathing ambient air. Lung auscultation revealed bibasal rhonchi. At presentation day (d) 4, SARS-CoV–2 was detected by real-time reverse transcriptase polymerase-chain-reaction (rRT-PCR) from a nasopharyngeal swab specimen. SARS-CoV–2 was again detected at d5–6 from nasopharyneal, sputum and faecal samples, but was undetectable from d7 (Fg.1a).. Blood C-reactive protein was elevated at 83.2, with normal lymphocyte counts (4.3x109/L [range 4.0–12.0x109/L]) and normal neutrophil counts (6.3x109/L [range 2.0–8.0x109/L]). No other respiratory pathogens were detected. Her management was intravenous fluid rehydration without supplemental oxygenation. No antibiotics, steroids or antiviral agents were administered. Chest radiography demonstrated bibasal infiltrates at d5, which cleared on d10 (Fig.1b), and she was discharged to home isolation on d11. Symptoms resolved completely by d13 and she remained well at d20 post-onset of symptoms. Progressive increase in plasma COVID–19-binding IgM/IgG antibodies, from d7 until d20 was observed (Fig.1c).
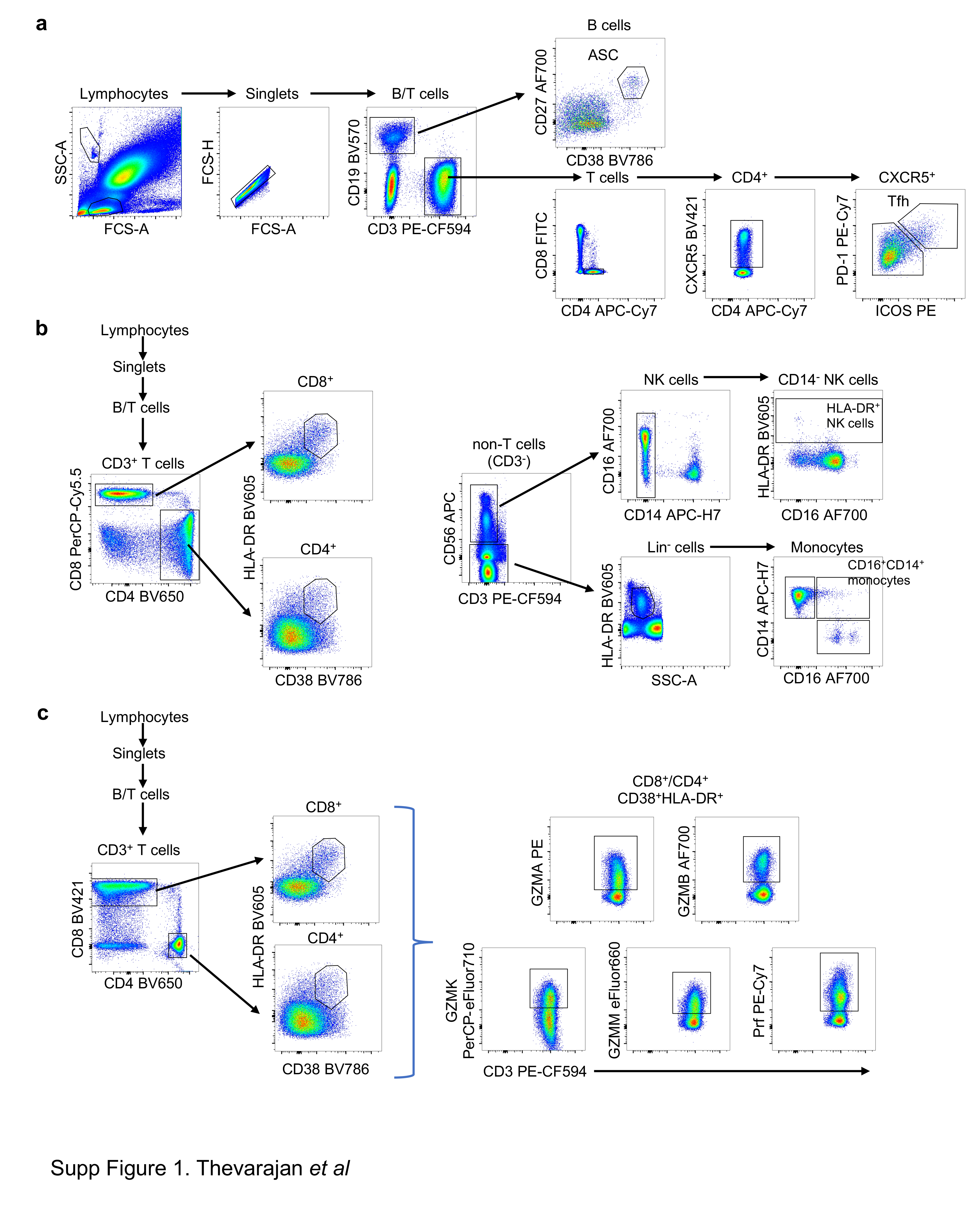
There are currently no data defining immune responses leading to viral clearance and clinical resolution of COVID–19. We addressed this knowledge gap by analysing the breadth of immune responses in blood prior to patient recovery. As antibody-secreting cells (ASCs) are key for the rapid production of antibodies following viral Ebola infection1,2, influenza virus infection and vaccination2,3; and activated circulating follicular T helper (cTfhs) are concomitantly induced following influenza vaccination3, we first determined the frequency of CD3-CD19+CD27hiCD38hi ASCs and CD4+CXCR5+ICOS+PD1+ cTfh responses at 3 days prior to symptomatic recovery. ASCs appeared in blood at the time of viral clearance at d7 (1.48%) and peaked on d8 (6.91%). Emergence of cTfhs occurred concurrently in blood at d7 (1.98%), with the frequency increasing on d8 (3.25%) and d9 (4.46%) (Fig.2a).. The peak of both ASCs and cTfhs was markedly higher in the COVID–19 patient than the baseline levels in healthy controls (average±SD: 0.61±0.40% and 1.83±0.77%, respectively, n = 5). Both ASCs and cTfhs were still prominently present at convalescence (d20) (4.54% and 7.14%, respectively; Fig.2a).. Our study provides evidence on the recruitment of both ASCs and cTfhs in patient’s blood whilst still unwell and 3 days prior to resolution of symptoms, indicating their importance in anti-viral immunity towards SARS-CoV–2.
Since co-expression of CD38 and HLA-DR is well defined as the key phenotype of CD8+ T-cell activation towards viral infections, we analyzed activation of CD8+ T-cells by CD38/HLA-DR co-expression. In accordance with previous reports on Ebola and influenza1,4, CD38+HLA-DR+ co-expression on CD8+ T-cells rapidly increased from d7 (3.57%) to d8 (5.32%) and d9 (11.8%), with a decrease at d20 (7.05%) (Fig.2b).. Furthermore, the frequency of CD38+HLA-DR+ co-expression on CD8+ T-cells in this patient was markedly higher than on CD8+ T-cells in healthy individuals (1.47±0.50%, n = 5). Similarly, CD38+HLA-DR+ co-expression increased on CD4+ T-cells between d7 (0.55%) and d9 (3.33%) in the patient, compared to healthy donors (0.63±0.28%, n = 5), although at lower levels than CD8+ T-cells. CD38+HLA-DR+ T cells, especially within CD8+ T-cells, produced higher amounts of granzymes A/B and perforin (~34–54% higher) than their parent (CD8+ or CD4+ populations, Fig.2b).. Thus, the emergence and rapid increase in activated CD38+HLA-DR+ T-cells, especially CD8+ T-cells, at d7–9 preceded resolution of symptoms.
We also analysed CD16+CD14+ monocytes, related to immunopathology, and activated HLA-DR+CD3-CD56+ NK cells (Fig.2c).. We detected reduced frequencies of CD16+CD14+ monocytes in peripheral blood at d7–9 (1.29%, 0.43%, 1.47%, respectively), compared to healthy controls (9.03±4.39%, n = 5). This might indicate efflux of CD16+CD14+ monocytes from blood to the site of infection, which remained low at d20 (2.24%). Low levels of activated HLA-DR+CD3-CD56+ NK cells were found in both the COVID–19 patient and healthy controls.
As high levels of pro-inflammatory cytokines/chemokines are predictive of severe clinical outcomes for influenza5, seventeen pro-inflammatory cytokines/chemokines were quantified in patient’s plasma. We found low levels of monocyte chemoattractant protein–1 (MCP–1; CCL2), important for the recruitment of monocytes, T-cells and dendritic cells to the site of infection (Fig.2d).. However, these MCP–1 levels were similar to healthy donors (22.15±13.81, n = 5), patients infected with influenza A (IAV) and influenza B viruses (IBV) at d7–9 (33.85±30.12, n = 5) and a patient with a known human coronavirus infection HCoV–229e (hCoV, 40.56). Substantial levels of RANTES (CCL5), involved in homing and migration of activated T-cells that express CCR5, were also detected in COVID–19 plasma but these were comparable to healthy donors (p = 0.412), IAV/IBV-infected patients (p = 0.310) and a hCoV-patient. Thus, in contrast to severe avian H7N9 disease with highly elevated IL–6 and IL–8, and intermediate IL–10, MIP–1β, IFN-γ5, minimal pro-inflammatory cytokines/chemokines were found in this patient with CoVID–19, even while symptomatic at d7–9.
Given that interferon-induced transmembrane protein–3 (IFITM3) single nucleotide polymorphism (SNP)-rs12252-C/C was linked to severe influenza5,6, we analysed the IFITM3- rs12252 SNP in this patient with COVID–19. Interestingly, the patient had the ‘risk’ IFITM3-rs12252-C/C variant (Fig.2e),, associated with clinical compromise for 2009-pH1N16 and severe/fatal avian H7N9 disease5. As the relative prevalence of IFITM3-rs12252-C/C risk variant in a healthy Chinese population is 26.5% (data from 1,000 genome project)5, further investigations of the IFITM3-rs12252-C/C allele in larger cohorts of patients with COVID–19 and its correlation with disease severity is worth pursuing.
Collectively, our study provides novel contributions to the understanding of the breadth of the immune response during a non-severe case of COVID–19. This patient did not experience complications of respiratory failure, acute respiratory distress syndrome, did not require supplemental oxygenation and was discharged within a week of hospitalization, consistent with non-severe but clearly symptomatic disease. We provide evidence on the recruitment of immune populations (antibody-secreting B cells, follicular T-cells, activated CD4+ and CD8+ T-cells), together with IgM-IgG SARS-CoV–2-binding antibodies, in patient’s blood prior to resolution of clinical symptoms. We propose that these immune parameters should be characterised in larger cohorts of patients with COVID–19 with different disease severity to understand whether they could be used to predict disease outcome and to evaluate new interventions to minimise severity and/or to inform protective vaccine candidates. Furthermore, our study indicates that robust multi-factorial immune responses can be elicited towards the newly-emerged SARS-CoV–2, and similar to the avian H7N9 disease7, early adaptive immune responses might correlate with better clinical outcomes.
{kind=link}