In the present MHRA approved study we investigated, for the first time, the quantitative and qualitative changes in gut microbial profiles of human volunteers with RMS prior to and following experimental infection with N. americanus, and following administration of anthelmintic treatment, and compared the findings with data obtained from an age- and gender-matched cohort of RMS patients subjected to placebo treatment.
Changes in gut microbial diversity
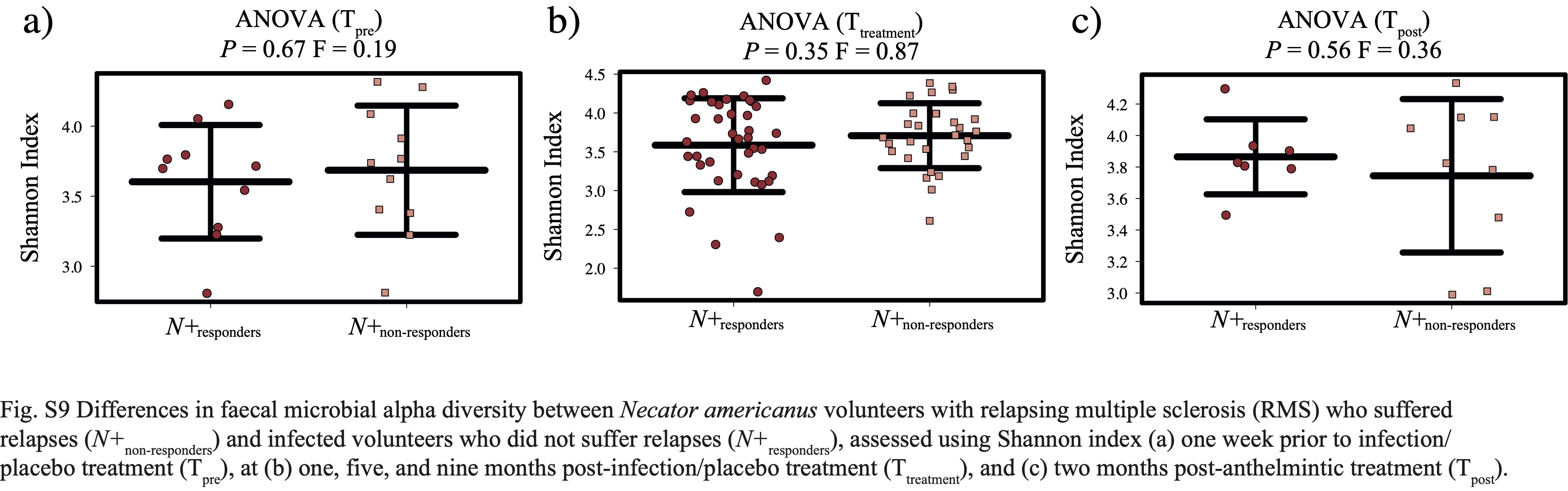
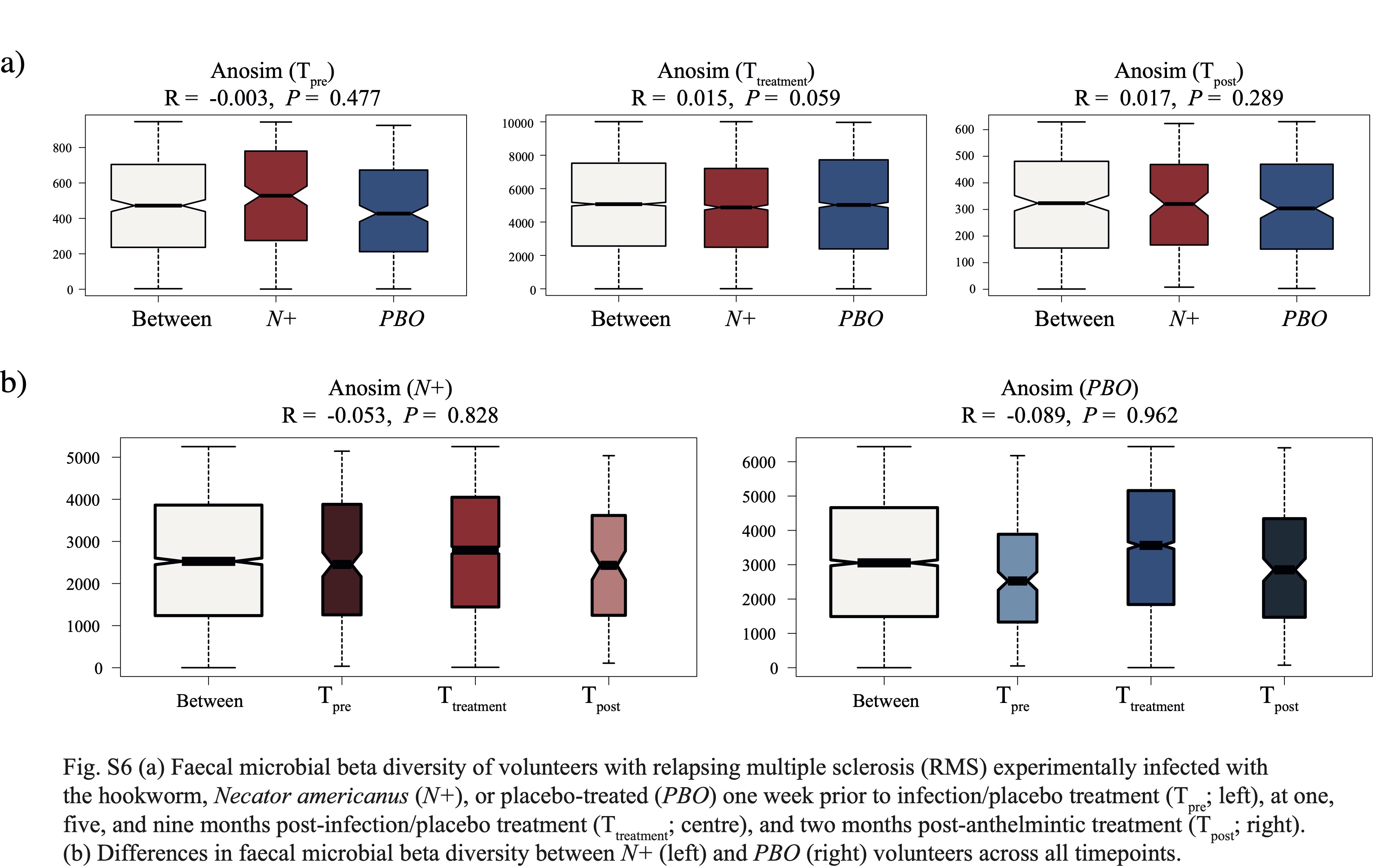
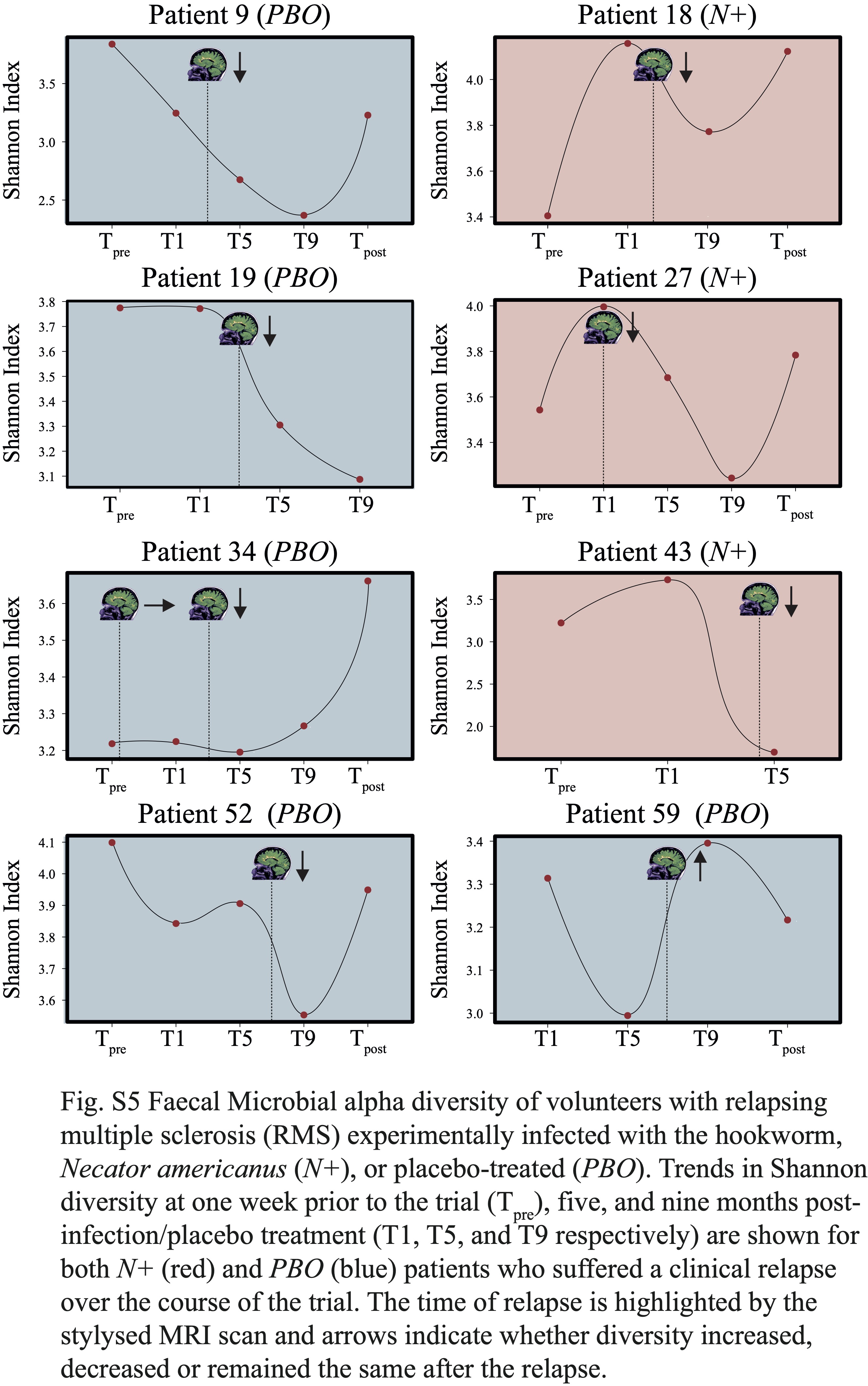
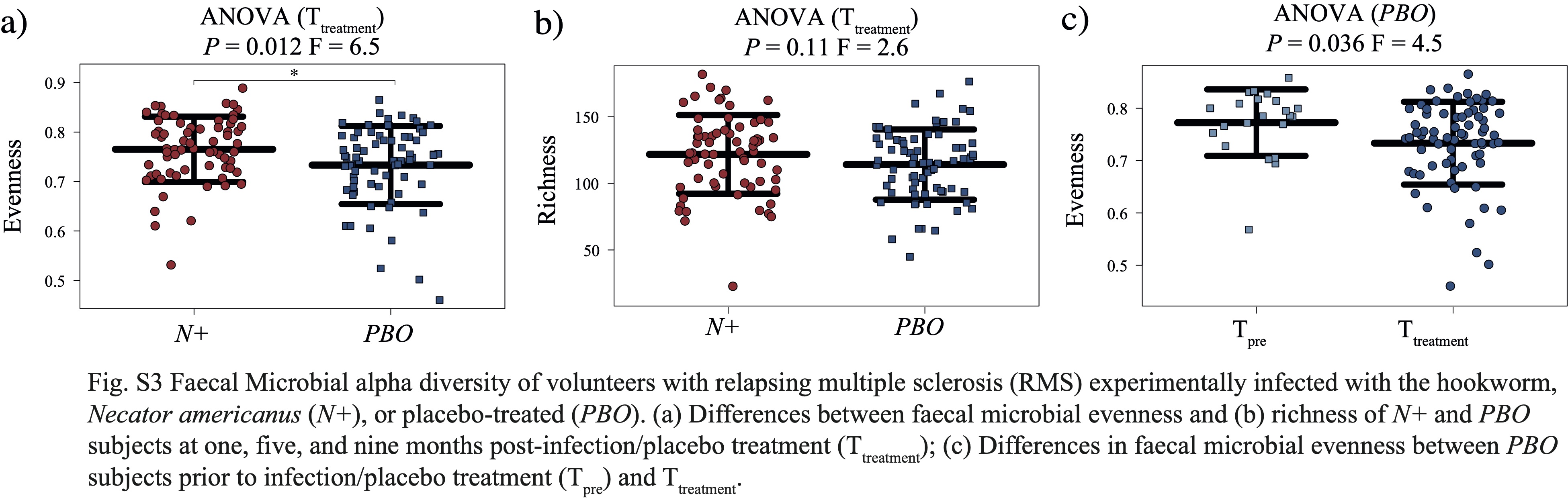
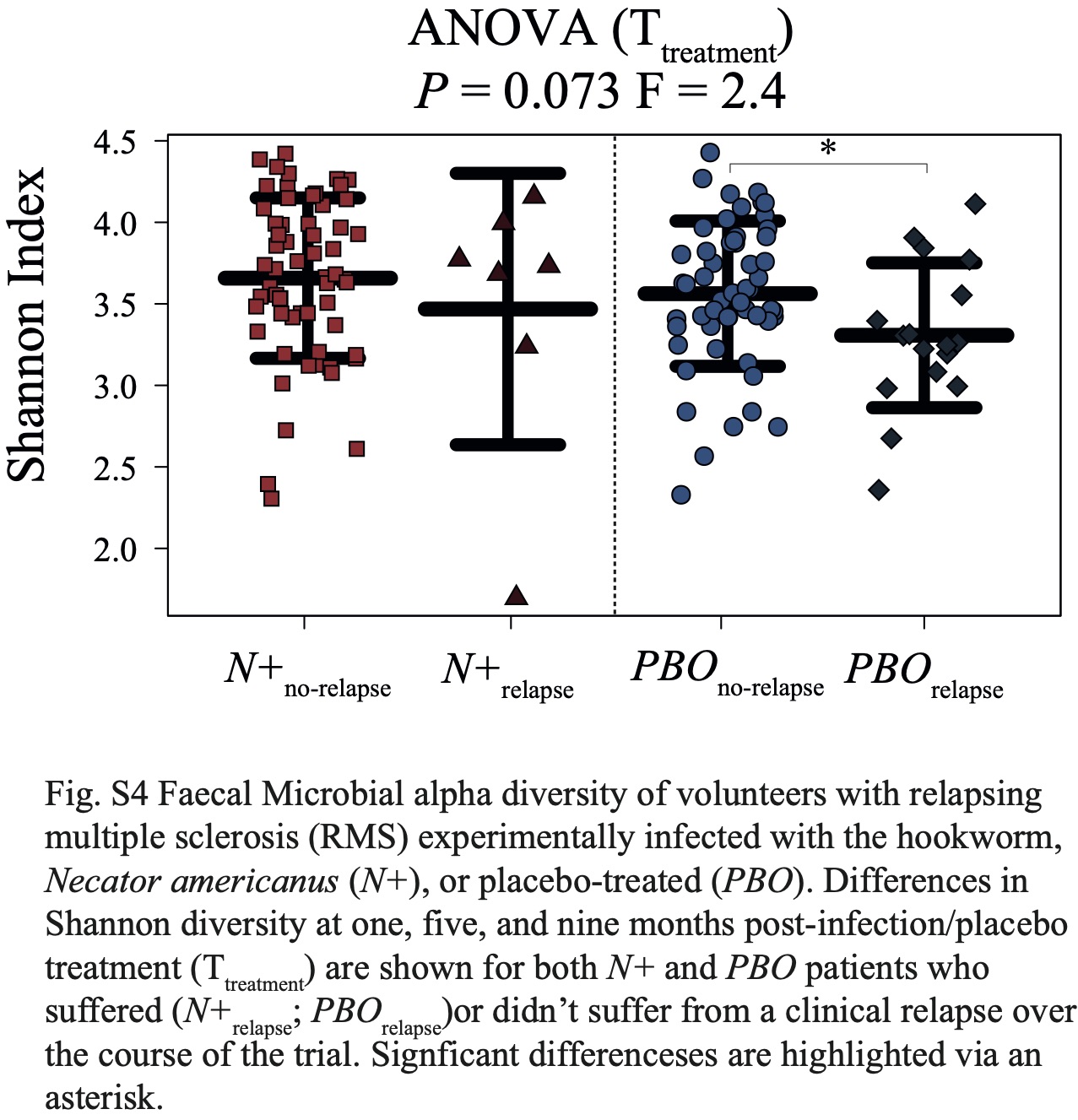
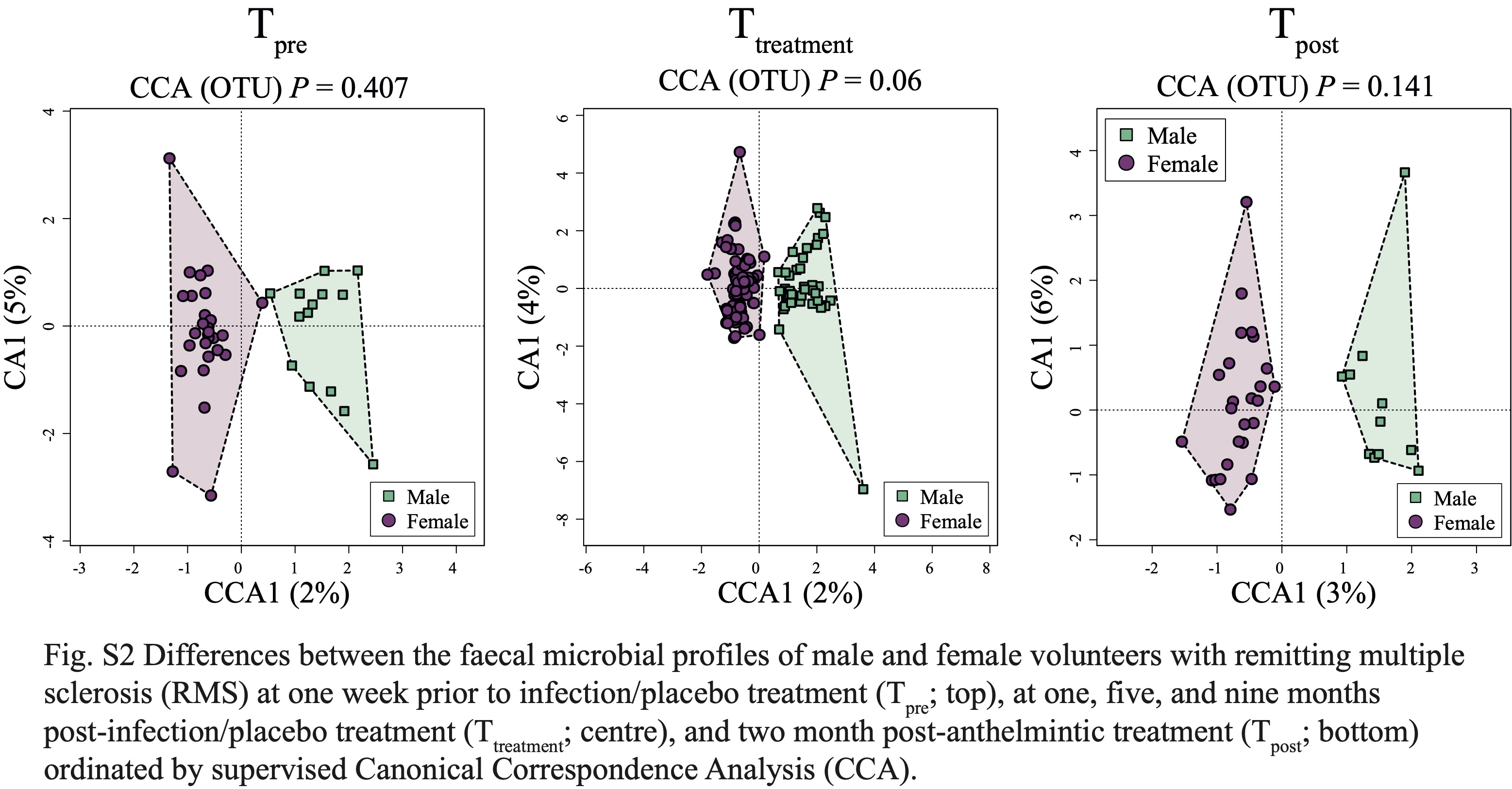
No substantial differences in gut microbiota composition were detected between N+ and PBO patients prior to experimental infection based on CCA analysis, while significant alterations in faecal microbial profiles were detected in the infected cohort compared with uninfected subjects post-worm colonisation. Of note, these differences were no longer significant post-anthelminthic treatment, thus suggesting a direct modulating effect of live hookworm infection on the microbial communities inhabiting the host gut. In particular, microbial alpha diversity was significantly higher in the gut of N+ subjects compared to PBO over the course of helminth treatment. It must however be pointed out that this observation was predominantly linked to a marked decrease of gut microbial alpha diversity, and specifically of bacterial evenness, in PBO over time. Parasite removal via anthelmintic treatment did not result in microbial alpha diversity alterations in both N+ and PBO. Notably, compared with values recorded prior to hookworm infection, this parameter was significantly increased in N+ subjects at end the of the trial, whilst the opposite trend was observed in the gut microbiota of PBO. This finding is of particular interest, since elevated levels of microbial alpha diversity are typically associated with a ‘healthier’ gut microbiome and overall host health [reviewed by 23]. Similarly, increases in gut microbial alpha diversity were reported in CeD subjects experimentally infected with N. americanus, which led the authors to speculate that this mechanism might be at least partially responsible for the therapeutic effect of deliberate helminth infections in individuals affected by selected allergic and autoimmune disorders [5, 9]. On the other hand, a decrease in gut microbial alpha diversity has previously been reported during MS relapses [24]. This matches our observations of a significantly lower microbial alpha diversity being detected at Ttreatment in the faecal microbiota of PBO volunteers who suffered relapses over the course of the trial, as well as decreased alpha diversity following specific relapse events in both N+relapse and PBOrelapse patients.
Gut microbial taxa expanded in the PBO cohort
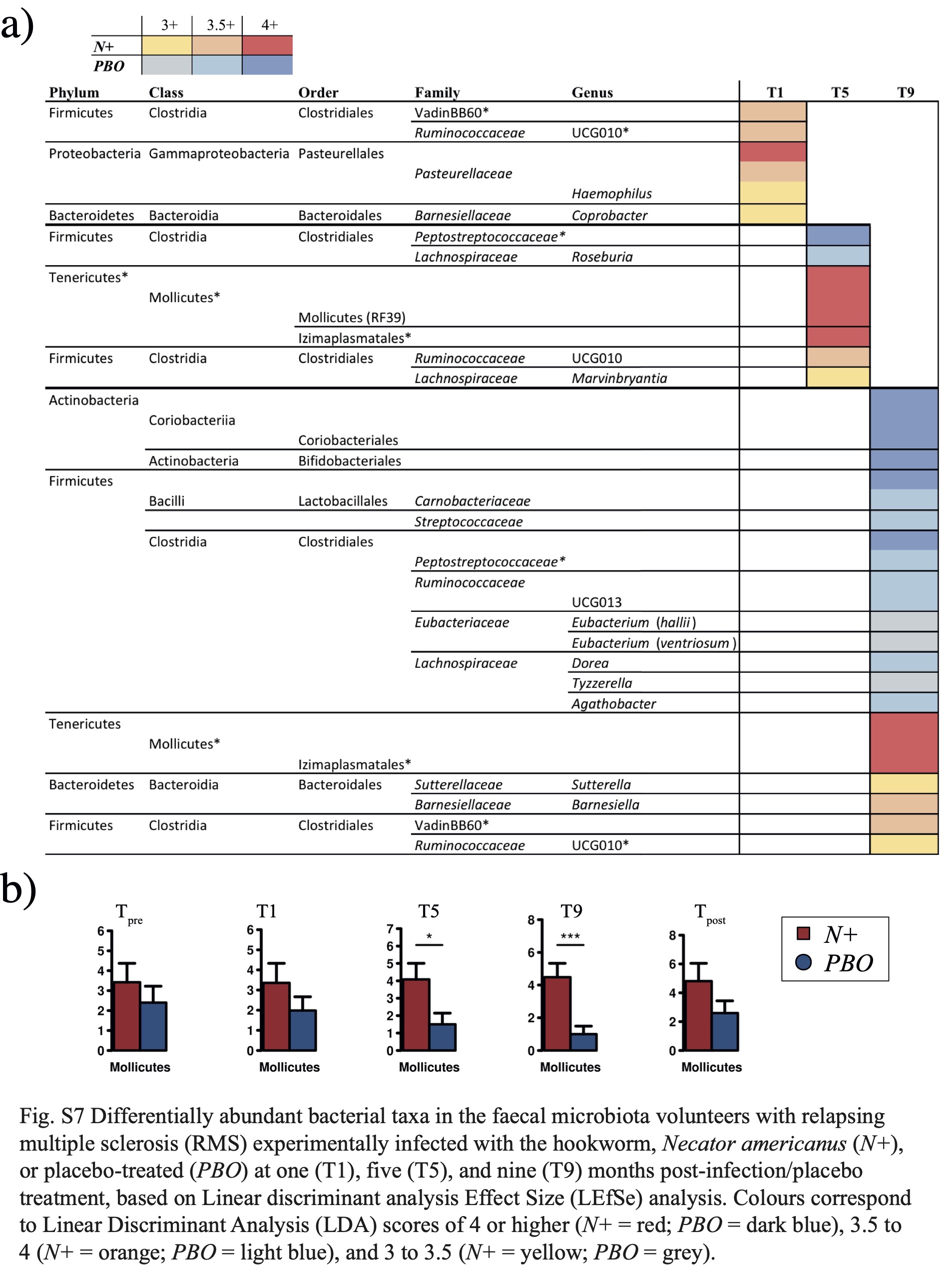
The relative abundances of several gut bacterial taxa were also significantly altered in N+ subjects compared with the PBO cohort, both prior to N. americanus experimental infections and, more markedly, post-helminth colonisation. Indeed, despite subject randomisation prior to the beginning of the study, we detected minor differences in gut microbial composition between study cohorts at Tpre. No prior characterisation of faecal microbial communities was conducted prior to group assignments; in addition, due to the intrinsic heterogeneity of gut microbial communities across any given population [25, 26], differences in the abundances of gut microbial taxa are frequently detected between groups of individuals enrolled in randomized, double-blinded, placebo-controlled trials [27, 28], with varying functional significance [28]. Nevertheless, in our study, most differences between N+ and PBO gut microbial profiles were detected post-infection, peaking at 9 months post-helminth colonisation. Notably, in PBO patients, bacterial taxa that have been previously associated with the gut microbiota of relapsing MS patients [24] were significantly expanded; in particular, a family of anaerobic bacteria, the Lachnospiraceae, including the genera Roseburia, Dorea, and Tyzzerella (amongst others), were significantly increased in the faecal microbiota of PBO compared to N+ subjects post-infection. Lachnospiraceae is a key family of the human gut microbiome that degrades complex polysaccharides to short-chain fatty acids (SCFAs), i.e. acetate, butyrate, and propionate, that are used for energy by the host. Notably, SCFAs are known for their anti-inflammatory properties [29]. This is of note, since Lachnospiraceae have been reported to be substantially expanded in the gut microbiota of individuals affected by pathological conditions, such as inflammatory bowel disease (IBD) [30]. In particular, the abundance of Dorea increases in the gut of irritable bowel syndrome patients [31], as well as during intestinal inflammation [32]. Whilst Dorea are known to produce the SCFA butyrate, this genus of bacteria also metabolises sialic acids, which are commonly found at the terminal ends of mucins; release of these acids is implicated in mucin degradation, with consequences ranging from increased gut permeability to compromised gut homeostasis; this cascade of events has been suggested to result in proinflammatory responses that promote chronic inflammation in MS [33, 34]. The expansion of Lachnospiraceae and its respective genera in the PBO cohort over the course of the trial is likely associated to MS disease progression and an immune shift towards a pro-inflammatory phenotype [34]. Of note, no significant expansion of Lachnospiraceae could be detected in the gut microbiota of N+ (as a whole) over time, with the exception of N+non-responders (see below).
Gut microbial taxa expanded in the N+ cohort
Bacteria belonging to the phylum Tenericutes were substantially and consistently increased in the gut microbiota of N+ patients post-infection. This phylum consists of the sole class Mollicutes, Gram‐ negative, small and wall-less bacteria that fulfil a diverse array of roles within the mammalian microbiome [reviewed by 35, 36]. The relative abundance of these bacteria has been reported to differ between the normal gut microbiota and that found in a wide range of autoimmune conditions, such as IBD [37, 38], Type 1 Diabetes [39-41], MS [42], and experimental autoimmune encephalomyelitis (EAE; a murine model of MS) [43]. Albeit inconsistently [40, 41], Tenericutes/Mollicutes are often reduced in the gut microbiota of the diseased cohort, when compared to healthy controls [37-39, 42, 43]. Notably, whilst one of these studies reported higher Tenericutes abundance in the gut microbiota of healthy controls than in paediatric MS patients, exposure to immunomodulatory drugs reversed this trend [42]. Indeed, these bacteria have been suggested to proliferate in Th2-dominant environments [8, 44]. Tenericutes/Mollicutes were also expended in the gut microbiota of vertebrates infected by Th2-inducing helminth parasites, human cohorts naturally infected with roundworms (i.e. Trichuris and/or Ascaris and/or hookworm) [45], rats infected with the tapeworm Hymenolepis diminuta [44], and primates with CID experimentally infected with T. trichiura [8]. This is in accordance with data from the WIRMS trial, that reported a markedly increased eosinophilia in N+ compared to PBO subjects [10]. Nevertheless, the functional consequences that expanded populations of Tenericutes/Mollicutes associated to infection by parasitic helminths play in the pathophysiology of the abovementioned chronic inflammatory and autoimmune disorders, as well as in RMS, remains to be determined. Alongside the evidence from previous studies, our data lends further support to the need for mechanistic investigations of the interactions between helminth and/or their products and this bacterial taxon in vitro and/or in vivo, to assist untangling its potential role in disease progression and prevention.
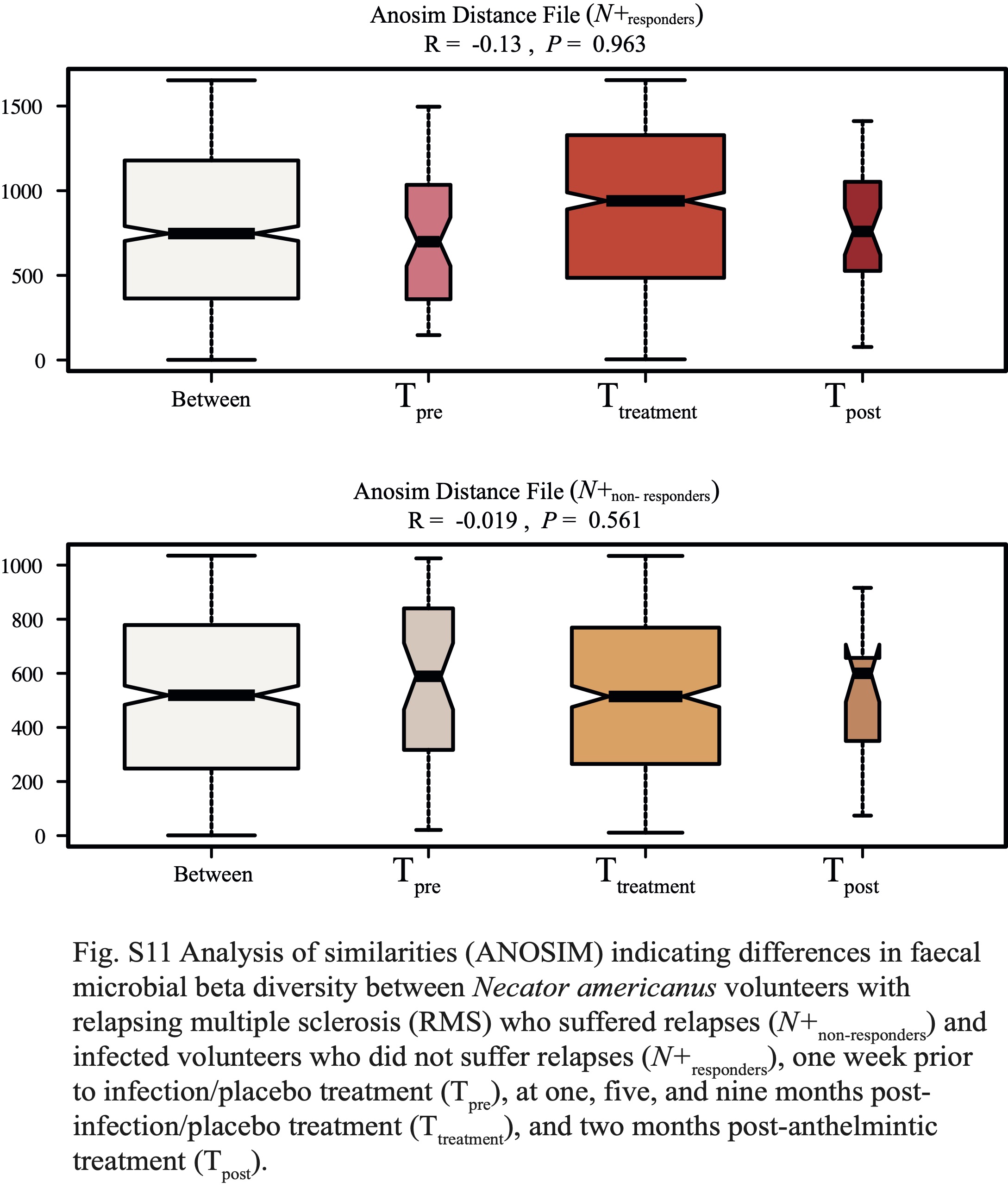
Gut microbial differences between N+responders and N+non-responders
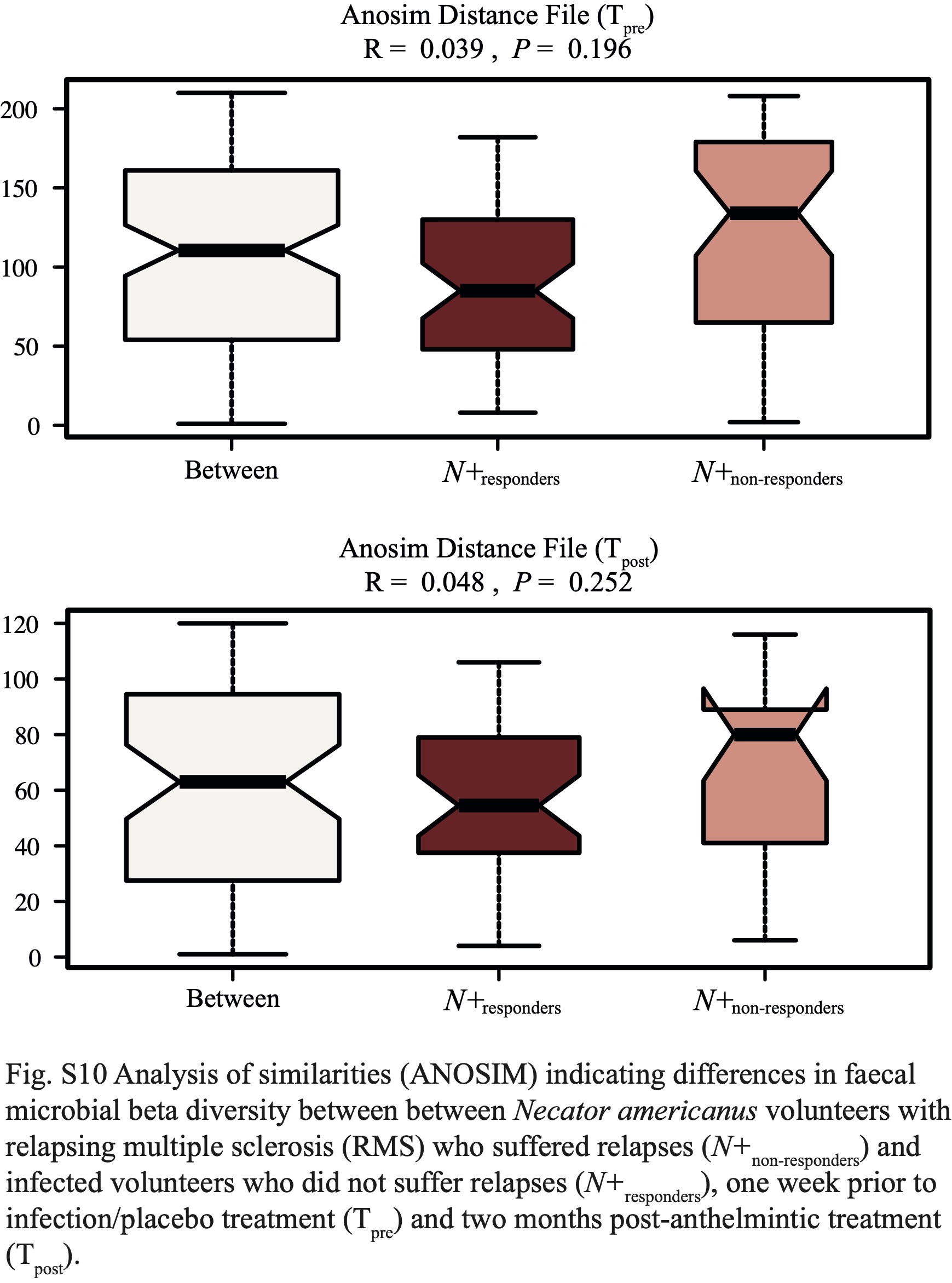
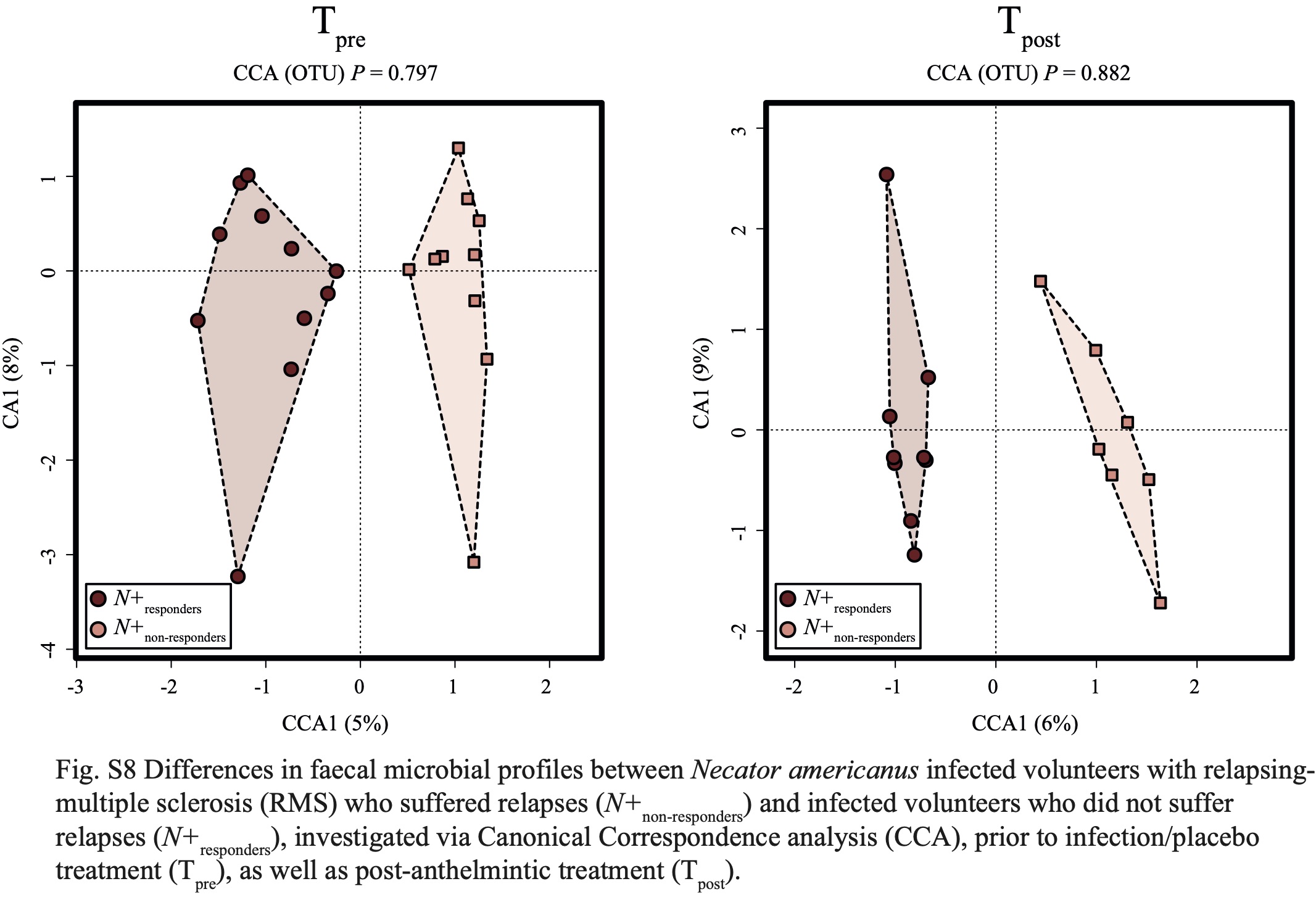
In addition, we investigated the differences in gut microbial composition between N+responders and N+non-responders, with the aim to identify potential bacterial candidates with roles in MS disease activity. Whilst the overall gut microbiota of these sub-cohorts did not differ substantially prior to helminth infection and or post-anthelmintic treatment, differences were recorded in the gut microbiota of these sub-groups post-helminth colonisation. In particular, while gut microbial alpha diversity remained similar between N+responders and N+non-responders, beta diversity was significantly higher in the former (albeit with a small effect size), indicating a ‘diversified’ gut microbiota in N+responders, either directly associated to N. americanus infection and/or, indirectly, to the immune responses mounted against the worms [1]. Meanwhile, the gut microbial beta diversity of N+non-responders was reduced over the course of the trial, possibly resulting from downstream effects of disease progression on the host gut microbiota [10, 46-48].
We detected significantly increased populations of Tenericutes/Mollicutes in the gut microbiota of N+responders, and expanded populations of Lachnospiraceae in N+non-responders, further supporting the hypothesis of a functional role of these taxa in the immune-modulatory properties of N. americanus. Furthermore, Flavobacteriaceae, the largest family in the phylum Bacteroidetes [49], was consistently increased in the gut microbiota of N+responders. This taxon has been repeatedly reported to be depleted in people suffering from autoimmune conditions, such as rheumatoid arthritis [50] and myasthenia gravis [51], although the functional importance of this taxon in the pathophysiology of these conditions remains elusive. Finally, in this study, we asked the question of whether some of the minor differences in gut microbial profiles observed between N+responders and N+non-responders prior to hookworm experimental infection might be associated with positive or negative clinical outcomes. Amongst others, the genus Parabacteroides was significantly more abundant in the gut microbiota of N+responders compared to N+non-responders. Notably, a recent study conducted in murine models of RMS and chronic-progressive MS identified Parabacteroides as more abundant in the gut microbiota of control mice [52]. Additionally, a previous study detected a significant reduction of Parabacteroides populations in the faecal microbiota of 71 MS patients not undergoing immunesuppressive treatment compared to that of 71 healthy control subjects [53]. Subsequent monocolonisation of antibiotic-treated mice with Parabacteroides distasonis led to significant increases in the CD4+IL-10+ T lymphocyte population in mesenteric lymph nodes and spleens [53]. Furthermore, stimulation of peripheral blood mononuclear cells (PBMCs) from MS patients or healthy controls with total bacterial extracts isolated from the stool samples of the same subjects resulted in the inability of PBMCs from MS patients to differentiate or expand CD25+FoxP3+ Treg populations [53]. This observation led the authors to hypothesise that prior exposure to P. distasonis or other “beneficial” bacteria may have contributed to the expanding regulatory T lymphocyte precursor populations in mice, hence promoting anti-inflammatory responses upon subsequent exposure to the same bacteria [53]. Furthermore, it is known that Parabacteroides metabolise phytoestrogens [54-56]. Estrogens have been shown to possess disease-suppressive properties in MS as evidenced in several studies in humans and animal models [57].
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}