Identification of a peptide that specifically binds to the extracellular domain of human VEGFR3.
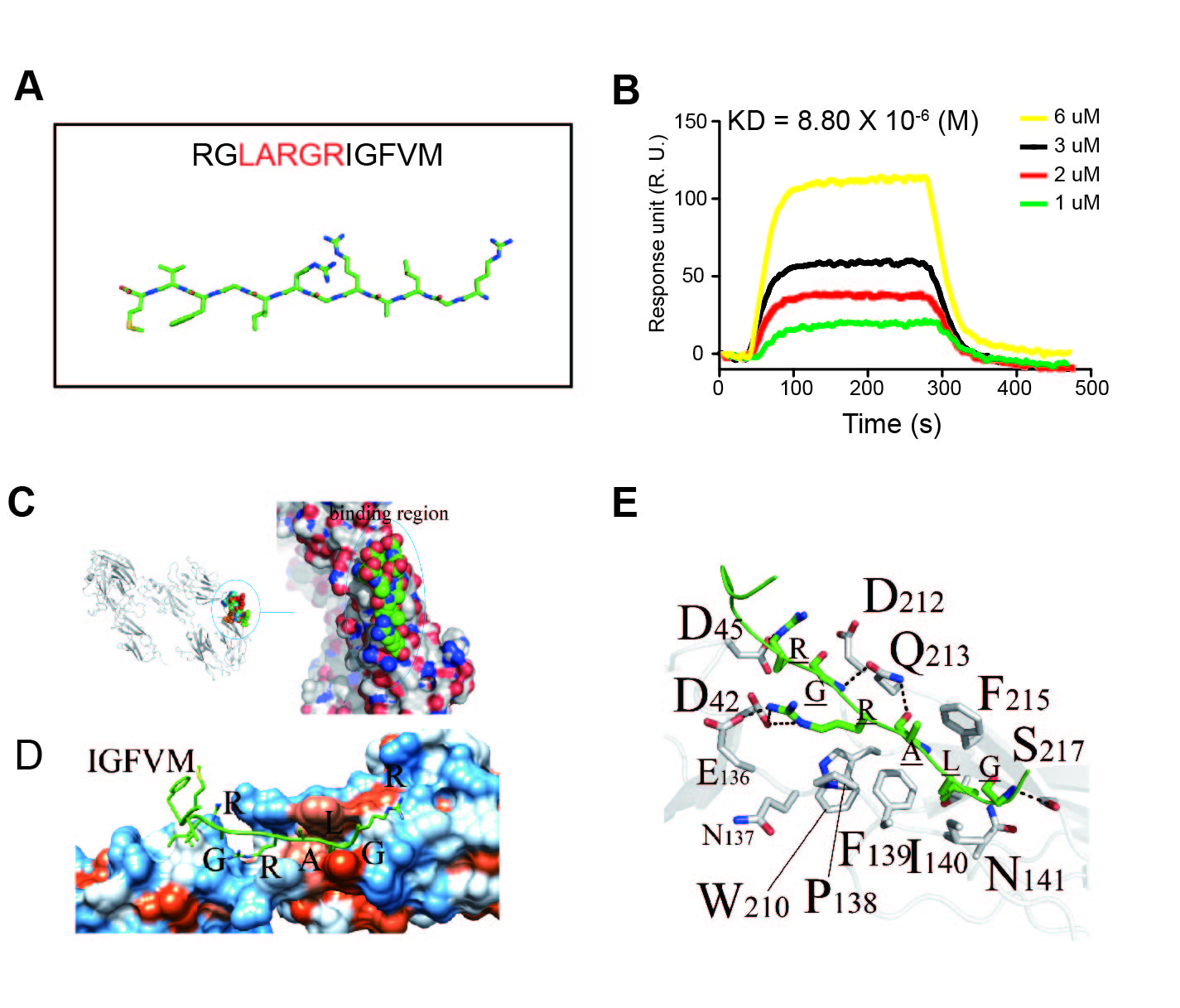
We use the FliTrx™ Random Peptide Display Library, which is based on a random 12-mer cyclic peptide display library, to screen the binding peptides with recombinant human VEGFR-3/Flt-4.After four rounds, 100 individual FliTrx clones were selected and their peptide-encoding inserts were sequenced and analyzed for potential repetitive peptide motifs. Analyzing the core amino acids by computerized analysis, one of the peptide sequences enriched in the selected clone pool was RGLARGRIGFVM (Fig S1 A). To further confirm the affinity of the selected peptide with VEGFR-3, RGLARGRIGFVM was synthesized and the SPR assay was utilized to measure the binding affinity. As shown in Fig S1 B, the RGLARGRIGFVM exhibited the binding kinetics with a Kd value of 8.80×10-6 (M).
To further analyze which sequence represents the core binding sequence of RGLARGRIGFVM with VEGFR3, we sought to identify the amino acid residues of VEGFR-3 that could be involved directly in the peptide-protein interaction. To this end, the homology model of VEGFR-3 was built using Modeller software, and then a molecular docking simulation between RGLARGRIGFVM and VEGFR-3 was performed. The docking region sites revealed that D42, E136, F139, F215, and D212 (VEGFR-3 D1-2) are the amino acids near the binding pocket (Fig. S1 E). The LARGR on the polypeptide chain is well attached to the binding pocket, and the affinity and hydrophobic effects are well-matched, but the polypeptide amino acids on both sides of RGLARGRIGFVM cannot be combined with the binding pocket, but are free in aqueous solution (Fig. S1C and D). The interaction energy between protein and Peptide 1 (RGLARGRIGFVM) was -23.10 kcal/mol.
To validate the results of computational protein structure modeling, LARGR was synthesized, and the SPR assay and the docking method was also conducted. Fig. 1C, D, and E showed that LARGR docked in the same region of VEGFR-3 D1-2 as RGLARGRIGFVM, and LARGR exhibited significant binding with VEGFR-3. The Kd value (2.87×10-5 M) was similar to the 12-mer peptide (Fig. 1B), and the interaction energy was -16.77 kcal/mol.
FITC-TMVP1 binds with VEGFR-3 in vitro
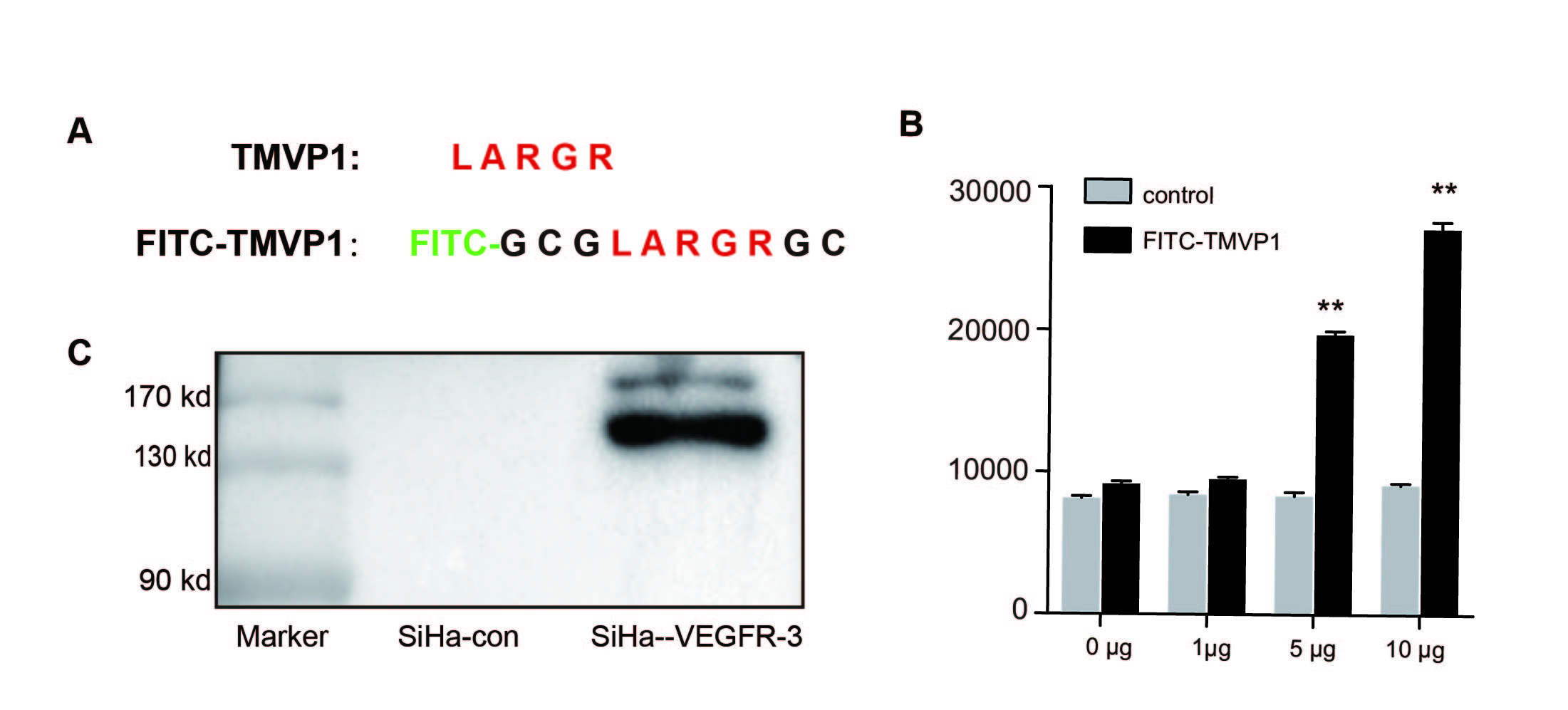
The immunofluorescence ELISA assay showed that the binding of FITC-TMVP1 to recombinant Human VEGF R3/Flt4 protein was FITC-TMVP1 concentration-dependent. When 5 or 10 µg of FITC-TMVP1 was added, the immunofluorescence signal density was significantly increased compared to that of the control group (**, P<0.01) as shown in Fig. S 2B.
LECs, HUVEC, and HK293 cells were incubated with 20 µM FITC-labelled TMVP1 at 37℃ for 60 min, the affinity of FITC-TMVP1 to cells presented that the LECs had an optimal binding and internalization ability(Fig. 2B). For further clarification of the VEGFR-3 targeting ability of TMVP1, we constructed VEGFR-3-overexpressed cells using the lentiviral vector, western blotting, and immunofluorescent assay detected the expression of VEGFR-3, as shown in Fig S2C. VEGFR-3 was abundantly expressed in SiHa cells. VEGFR-3 over-expressed cells and control SiHa cells were incubated with 20 µM FITC-conjugated synthetic TMVP1 and VEGFR-3 antibody. Cell nuclei were stained with DAPI. The fluorescent images in the tumor cell were examined using laser confocal microscopy. The results showed that the location of VEGFR-3 staining (red fluorescence) in the cell membrane was coincident with TMVP1 green fluorescence. Conversely, FITC-TMVP1 did not bind in the VEGFR-3-negative cells(Figure 2B).
FITC-TMVP1 targets lymphatic vessels in vitro and in vivo
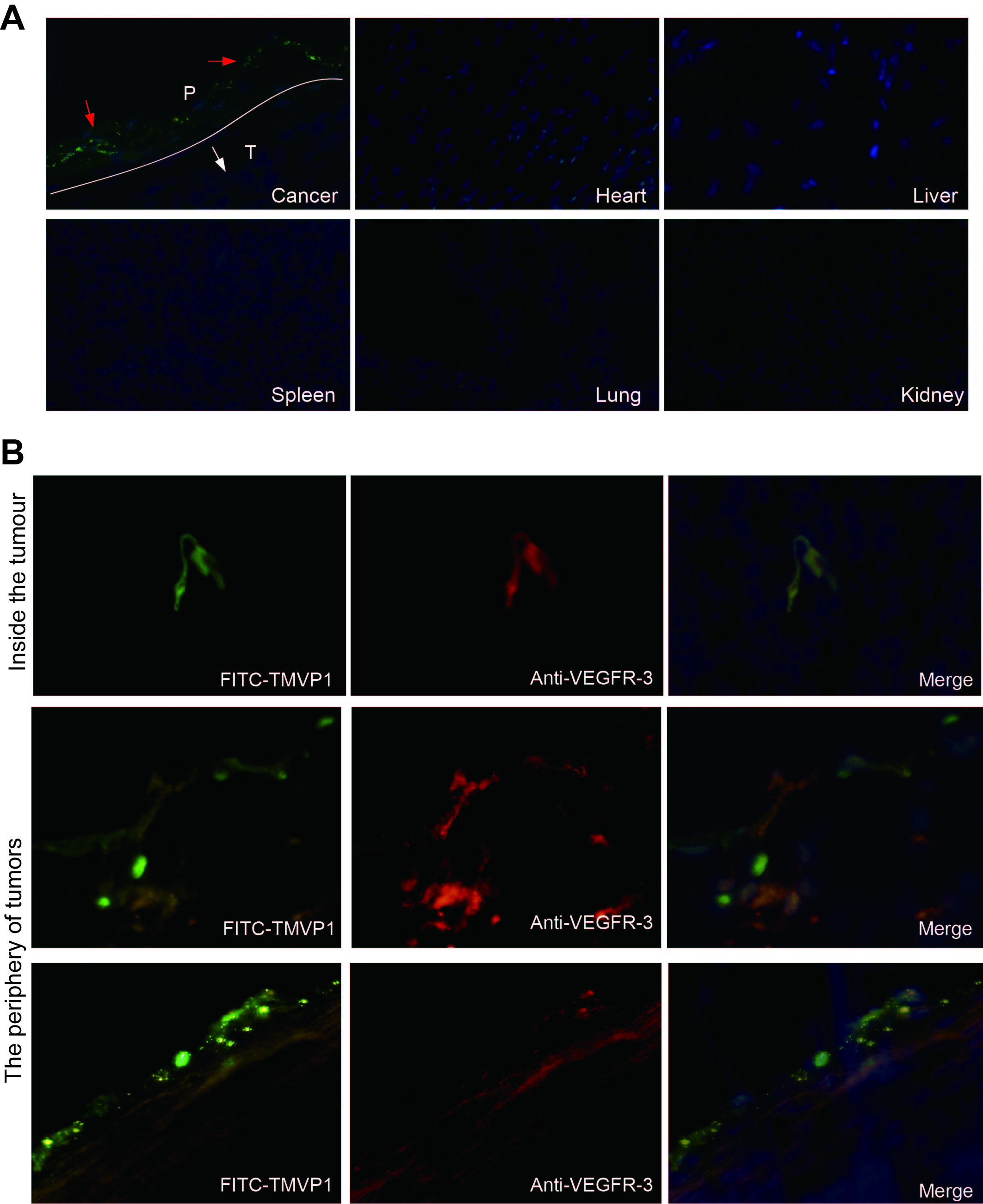
To determine whether TMVP1 could target tumor lymphatic vessels in vivo, the corneal micropocket mouse 4T1 tumor model was established. As shown in Fig. 3A, 1 h after i.v. injection, the strong green fluorescence accumulated within the micro-bag cornea of tumor model, while only background fluorescence was observed in normal control Figure. At the same time, the tumor frozen sections were immunostained using VEGFR-3 antibody, the results showed that the VEGFR-3 staining (red fluorescence) in the vasculature was coincident with TMVP1 green fluorescence (Fig. 3B). The biodistribution of FITC-TMVP1 was mainly accumulated at the periphery of growing tumors (Fig. S3 A), in accordance with corneal micropocket mouse 4T1 tumor model, the VEGFR-3 staining (red fluorescence) in the vasculature was also coincident with TMVP1 green fluorescence in the 4T1 subcutaneous tumor model (Fig. S3 B).
The clinical tissue samples from breast cancer patients were dissected to study the binding specificity of TMVP1with neoplastic lymphatic vessels. A total of 12 human breast cancer specimens were analyzed in this study. The FITC-TMVP1 staining was observed to be completely coincident with anti-VEGFR-3, and partially colocalized with anti-LYVE1 staining with in all the 12 breast tumor sample (Fig. 3C).
TMVP1 targets tumor tissues as indicated by 99mTc labeling
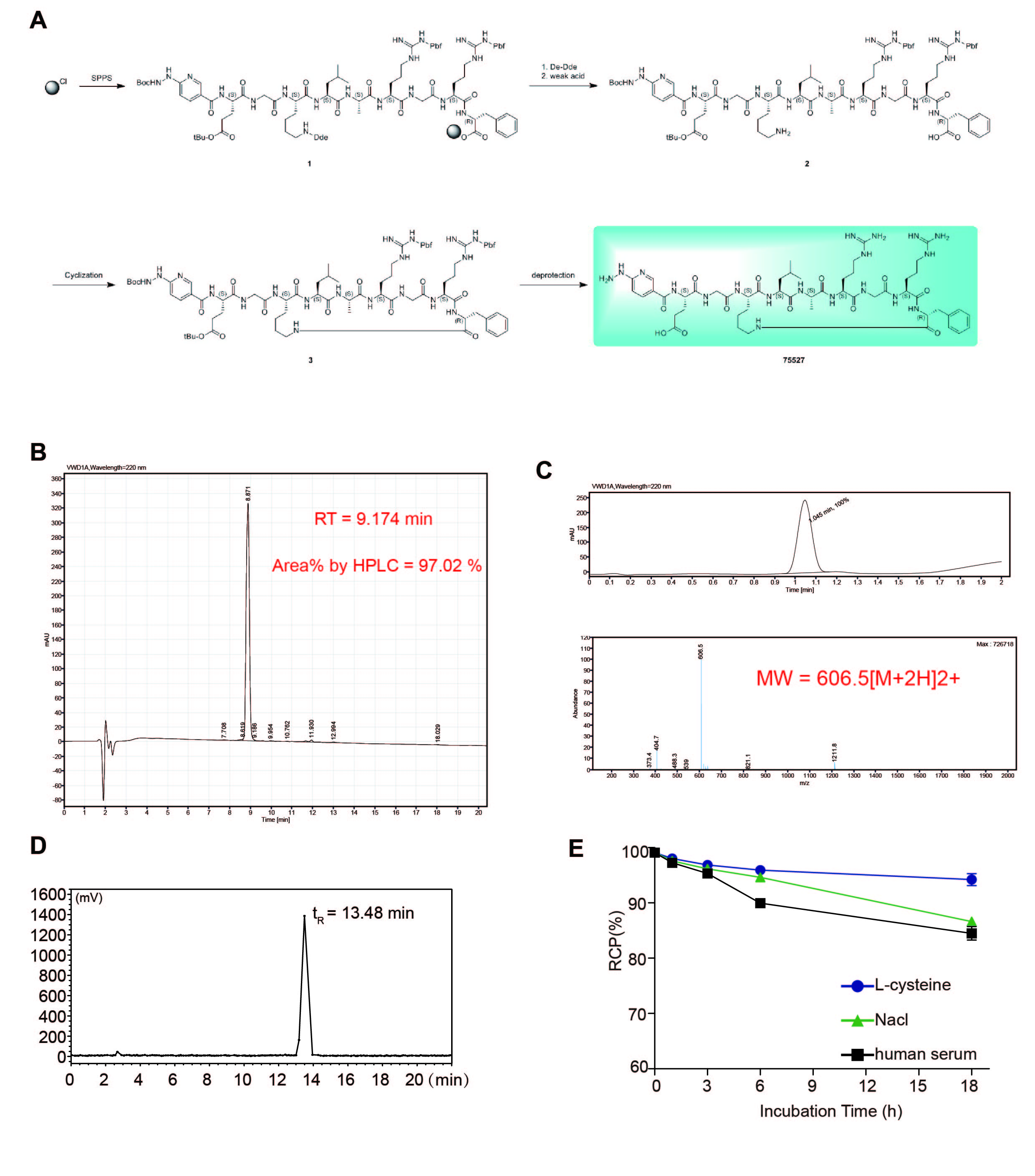
The Kd value of HYNIC-TMVP1 with VEGFR-3 was 2.96×10-6 M. The SPR assay verified that the TMVP1 binding affinity with VEGFR-3 was not reduced (Fig 4B). Then HYNIC-TMVP1 was labeled with 99mTc, and the radiochemical purity of 99mTc-HYNIC/EDDA-TMVP1 was measured by radio-HPLC and ITLC-SG, the retention time was 13.48 min (Fig. S4 D), the radiochemical purity was greater than 95%. The solution stability of 99mTc-HYNIC/EDDA-TMVP1 was monitored by ITLC-SG in saline, in serum, and the presence of excess cysteine, and the radiotracer was stable in the all these solutions (Fig. S4 E).
Fig. 4C illustrates representative SPECT images of tumor-bearing mice at 30 min, 1 h, and 2 h after administration of ∼ 18.5 MBq of 99mTc-labeled TMVP1 with or without excess TMVP1 pre-injection. The SPECT images showed that the B16 and 4T1 tumors were clearly visualized after administration, but SiHa tumors were poorly visualized. Meanwhile, the tumor imaging could be blocked by co-injection of excess cold TMVP1 (Fig. 4C and D). In addition, the radiotracer accumulated mainly in the kidney and bladder, which reflected the renal-mediated clearance of the radiotracer. IHC revealed VEGFR-3 expression in the above three tumor tissues. The results showed that the VEGFR-3 expression was in accordance with the accumulation of 99mTc-labeled TMVP1 (Fig. 4E).
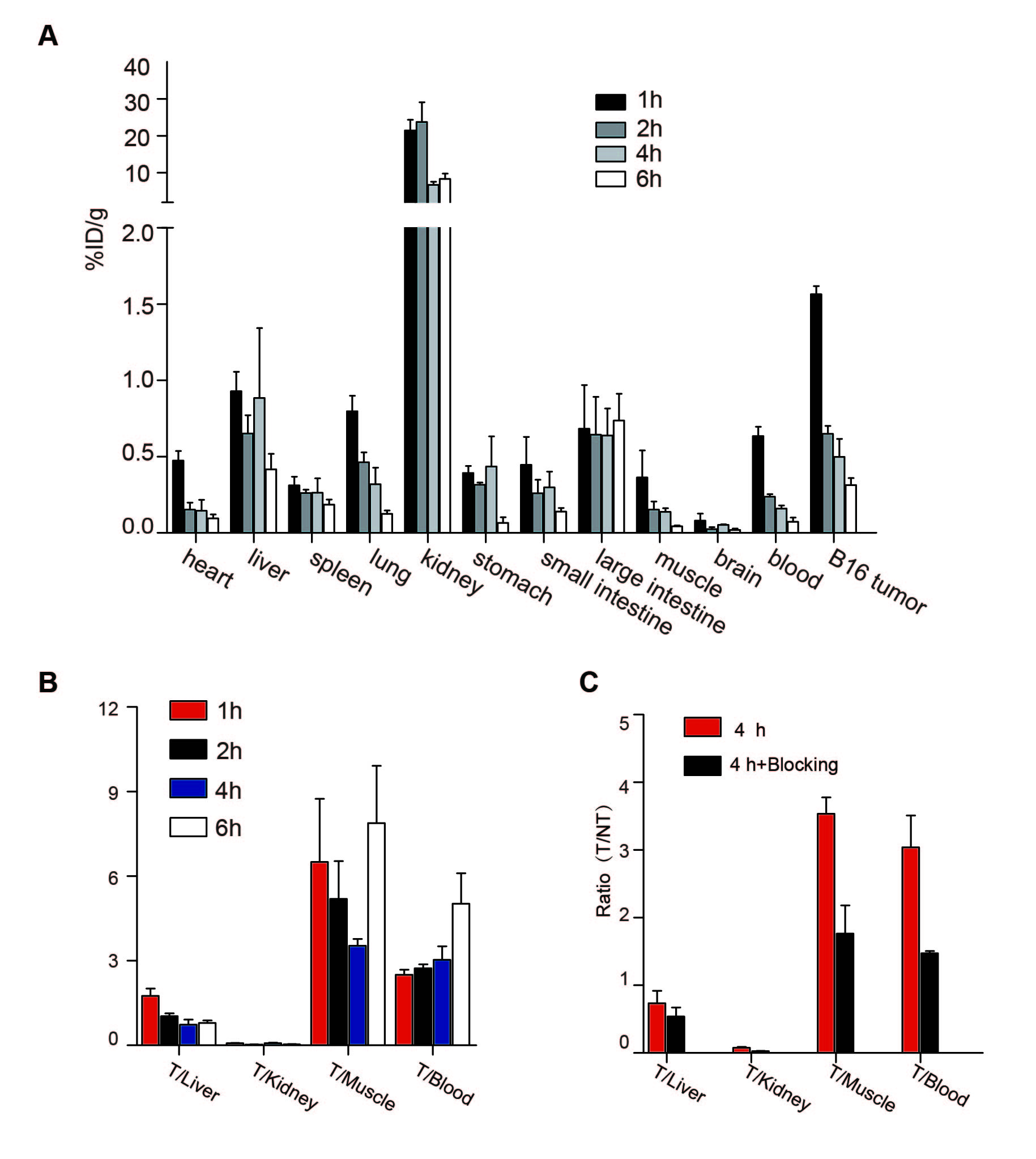
To further understand the biodistribution characteristics of 99mTc-labeled TMVP1, nude mice bearing 4T1 xenografts were used to evaluate the biodistribution characteristics and excretion kinetics. Fig. S5A presents the %ID/g organ uptake for the tracer. In general, the radiotracer tumor uptake was 0.65 ± 0.09, and 0.314 ± 0.08 %ID/g at 2 h and 4 h, respectively, with extremely short blood retention times. The tumor/blood ratios were 2.74 ±0.24 and 5.02 ±1.88 at 2 h and 4 h p.i., respectively (Fig. S5 B). The blocking experiment demonstrated that the accumulation of radiotracer could be significantly reduced by pre-injection of excess TMVP1 (Fig. S5 C).
TMVP1-KLA inhibits LECs to form lymphatic-like tube in vitro and in vivo. Three-dimensional visualization of tube-formation and the in vitro sprout forming assay were performed, and frank tube-like structures were observed in lymphatic endothelial cells, and their formation was enhanced by cytokine rhVEGF-C. When TMVP1-KLA or TMVP1-KLA plus rhVEGF-C were added, the numbers of lymphatic sprouts and tube-like structures were significantly reduced compared to the control groups, and some cells formed spherical apoptotic cells (Fig. 5A, B). The percentage of the average sprout length in TMVP1-KLA + rhVEGF-C- and TMVP1-KLA-treated LECs was, respectively, 5.2 ±1.1 % and 4.3+1.0 % significantly lower than that of rhVEGF-C-treated or control LECs. The percentage of tube-like structures coverage area in TMVP1-KLA + rhVEGF-C- and TMVP1-KLA-treated LECs were, respectively, 5.8 ±0.5 %, and 2.6 ± 0.4 %, which was significantly lower than that of the control.
To reveal the TMVP1-KLA inhibition of lymphangiogenesis, we chose the murine corneal transplantation model for our study. The number of lymphatic vessels found in the peptide-treated group was significantly less than that of the control group, as shown in Fig. 5C and D. Few lymphatic vessels were observed in the TMVP1-KLA treated group. The area of the LYVE1-positive staining in the TMVP1-KLA-treated group was 12.3 ±1.2 %, much less than that of the control group (p<0.01) (Fig. 5D). The arrangement of the tissue nuclei after TMVP1-KLA treatment was observed, with tightly packed nuclei and less lumen distribution. However, tissues in the control group were relatively loose, with more lumen structures.
TMTP1-KLA inhibits tumor lymphangiogenesis and development in vivo.
We next sought to explore whether the TMVP1-KLA peptide might represent a possible therapeutic strategy to suppress tumor lymphangiogenesis in vivo. We analyzed the effects of TMVP1-KLA on MDA-MB-231-derived orthotopic breast cancer lymphangiogenesis in mice. As shown in Fig. 6 A-C, TMVP1-KLA was observed to inhibit orthotopic breast tumor growth effectively. The mean tumor volumes of TMVP1-KLA-treated mice were 0.24 mm3, which were significantly (P < 0.05) smaller than that of TMVP1-treated mice (0.87 ±0.07 mm3; Fig. 6B). The tumor weight from the TMVP1-KLA group was 1.2 ±0.5 g, which was also significantly lower than that of the control group (Fig. 6C). Thus, TMVP1-KLA could significantly suppress tumor growth in vivo unlike TMTP1.
The tumor lymphangiogenesis vessel number of the mouse tumor tissue was observed between two groups using immunohistochemical staining. The area of LYVE1-positive staining decreased significantly in the TMTP1-KLA peptide-treated group compared to that in the control group (Figure 6D, E). Thus, these results showed a vigorous anti-lymphangiogenic effect of TMTP1-KLA peptide in vivo.
Effects of TMTP1-KLA on the inhibition of metastatic tumor cell extravasation into the lungs
An experimental lung metastasis model was applied to determine whether the TMTP1-KLA inhibition of metastasis was specifically due to decreased lymphatic vessels. In the TMVP1-KLA-treatment group, lung alveolar tissue and decreased numbers of cancer nests were observed, while in the control group, metastatic invasion left little alveolar lung tissue, as shown by HE staining. TMVP1-KLA effectively inhibited 4T1 subcutaneous tumor growth. The mean tumor volumes of TMVP1-KLA-treated mice were significantly (P < 0.05) smaller than those of control mice (as shown in Fig S6B). There were no deaths observed in mice treated with TMVP1-KLA.
Moreover, the lungs of TMVP1-KLA-treated mice showed significantly smaller volumes and fewer micro-metastases than that of control mice (Fig. S 6A, C; P <0.01). Histologically, TMVP1-KLA–treated mice showed clumps of metastatic cells lodged within lymphatic vessels that failed to breach the lymphatic basement membrane and enter the lung parenchyma (Fig. S 6A) (significantly more visible lung metastases were visualized in the control group; P <0.01). These experimental results showed that TMVP1-KLA could inhibit breast cancer growth and metastasis.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}