IGF2BP2 mainly combined with SUMO1 to form SUMO modification
In order to determine whether IGF2BP2 can be SUMOylated in cells, HA-IGF2BP2, His-SUMO1, His-SUMO2, His-SUMO3 and Flag-Ubc9 were respectively transfected into HEK293T cells. Ni2+-NTA agarose bead pull-down assays were used to pull down the His-SUMO conjugates for Western blotting. Our results showed that IGF2BP2 could be modified by SUMO1, SUMO2 and SUMO3, and the modification by SUMO1 was the strongest (Fig. 1A). Therefore, the follow-up studies focused on the modification effect between IGF2BP2 and SUMO1.
SENP1 is one of the sumo specific proteases and participates in the process of SUMO modification. This study continues to verify the effect of SENP1 on SUMOylation of IGF2BP2. We transfected HA-IGF2BP2, His-SUMO1, Flag-Ubc9 and myc-SENP1 into HEK293T cells, respectively. Ni2+-NTA agarose bead pull-down assays were used to verify if SENP1 can remove the SUMO1 modification. We found that Ubc9 significantly increased the SUMO1 modification of IGF2BP2, while SENP1 significantly decreased the modification effect above, which played a role of de-SUMOylation (Fig. 1B).
In addition, in this study, plasmid of SENP1 knockdown was stably transferred to HEK293T cells, and the SUMO1 modification of IGF2BP2 was further verified by Ni2+-NTA agarose bead pull-down assays. Compared with NC group, the SUMOylation of IGF2BP2 in SENP1 (-) group was significantly enhanced (Fig. 1C).
His-SUMO1, Flag-Ubc9 with or without SENP1 were co-transfected into HEK293T cells. Ni2+-NTA agarose bead pull-down assays were used to verify whether endogenous IGF2BP2 can combined with SUMO1 and be SUMOylated by SUMO1. The SUMOylation bands were detected by using anti-IGF2BP2 antibody, and SUMO1 modification was the best in the presence of Flag-Ubc9, but could almost be removed by SENP1 (Fig. 1D).
In this study, co-immunoprecipitation (CO-IP) was used to further verify the binding and modification of endogenous SUMO1 on endogenous IGF2BP2. First, HEK293T cells were stably transfected with knockdown SENP1, knockdown Ubc9 and sh-NC plasmid respectively, and then, the lysates were immunoprecipitated with SUMO1, IGF2BP2 or normal IgG antibodies. Finally, western blot was performed with anti-IGF2BP2 and anti-SUMO1 antibodies. The results showed that the endogenous SUMO1 could bind and modify the endogenous IGF2BP2 in HEK293T cells. The SUMOylation of IGF2BP2 was enhanced by SENP1 knockdown, while the SUMOylation of IGF2BP2 was significantly inhibited byUbc9 knockdown. Compared with control group, input group confirmed that the knockdown ofSENP1 and Ubc9 did not affect IGF2BP2 expression (Fig. 1E).
To further deepen understanding the mutual modification of endogenous IGF2BP2 and endogenous SUMO1 in cells. Lysates of U87 and U251 cells were used for IP with anti-SUMO1, anti-IGF2BP2 and anti-IgG antibodies, and then western blot was performed with anti-IGF2BP2 and anti-SUMO1 antibodies. The results indicated that endogenous IGF2BP2 could bind to endogenous SUMO1 in glioma cells (U87 and U251) and undergo SUMOylation (Fig. 1F-G)
K497, k505 and k509 are the main SUMO-sites of IGF2BP2
In order to determine the main SUMO-sites of IGF2BP2 in human, we predicted that lysines (Ks) at K497, k505 and k509 could be mutated to arginine (R) according to the analysis results of bioinformatics software, and then as the main SUMO-sites for SUMOylation identification (Fig. 1H). we mutated the predicted lysins of IGF2BP2 protein, then co-transfected three single-lysin mutants and triple-lysin mutant with plasmids His-SUMO1 and Flag-Ubc9 into HEK293T cell for SUMO modification analysis by Ni2+-NTA agarose bead pull-down assays. The results indicated that compared with the wild-type (-Wt), the SUMO modification levels of IGF2BP2 were decreased in K497R, K505R, K509R and 3KR (K497/505/509R), and the SUMO modification level decreased most significantly in 3KR (K497/505/509R) (Fig. 1I).
SUMO modification enhanced the stability of IGF2BP2, but did not affect its subcellular localization
This study confirmed that SUMO modification could enhance the stability of IGF2BP2 by blocking the degradation process of IGF2BP2 in cells. First, the study found that there was no significant difference in the mRNA expression levels of IGF2BP2-Wt and IGF2BP2-3KR in HEK293T cells, but the protein expression level of IGF2BP2-3KR was significantly lower than that of IGF2BP2-Wt (Fig. 2A-B). Second, we co-transfected HA-IGF2BP2-Wt, HA-IGF2BP2-3KR, His-SUMO1 and myc-SENP1 into HEK293T cells, and used cycloheximide (CHX) to inhibit the synthesis of de novo proteins for12 hours. We found that compared with the HA-IGF2BP2-Wt group alone, the expression level of IGF2BP2 protein in HA-IGF2BP2-Wt and His-SUMO1co-transfected cells was significantly increased, but the protein expression enhancement effect was abolishedwhen co-transfected with myc-SENP1or HA-IGF2BP2-3KR (Fig. 2C). It is indicated that the degradation of IGF2BP2 was slowed down when conjugated with SUMO1. Third, the degradation of IGF2BP2 mainly occurs through ubiquitin proteasome pathway. Compared with chloroquine (lysosomal inhibitor), MG132 (proteasome inhibitor) significantly blocked IGF2BP2 degradation in the presence of CHX (Fig. 2D). Moreover, the degradation time of HA-IGF2BP2-3KR group was shorter than that of HA-IGF2BP2-wt group, and chloroquine did not block the degradation, while MG132 significantly blocked the degradation (Fig. 2D). Based on the above results, SUMOylation can effectively protect the substrate protein IGF2BP2 from degradation by ubiquitin proteasome pathway, thereby enhancing its stability.
In order to estimateif SUMOylation affects the subcellular localization of IGF2BP2, we co-transfected HA-IGF2BP2, His-SUMO1, Flag-Ubc9 and myc-SENP1 into HEK293T cells, and then the cytoplasmic and nuclear proteins were separated and extracted. Finally, the expression of IGF2BP2 protein was detected by western blot. The results showed that IGF2BP2 was mainly located in the cytoplasm, and SUMOylation did not affect the subcellular localization of IGF2BP2 (Fig. 2E).In this study, IGF2BP2-Wt and IGF2BP2-3KR were transfected into U87 and U251 cells, and the effect of SUMO-sites mutation on the subcellular localization of IGF2BP2 was detected by cytoplasmic and nuclear fractionation and Immunofluorescence staining. The results showed that after transfection of IGF2BP2-Wt and IGF2BP2-3KR, IGF2BP2 was still mainly located in the cytoplasm (Fig. 2H-G).
IGF2BP2 and OIP5-AS1 were highly expressed in both glioma tissues and cells. The SUMO-site mutates (IGF2BP2-3KR), knockdown of IGF2BP2 and OIP5-AS1 inhibited the VM of glioma cells
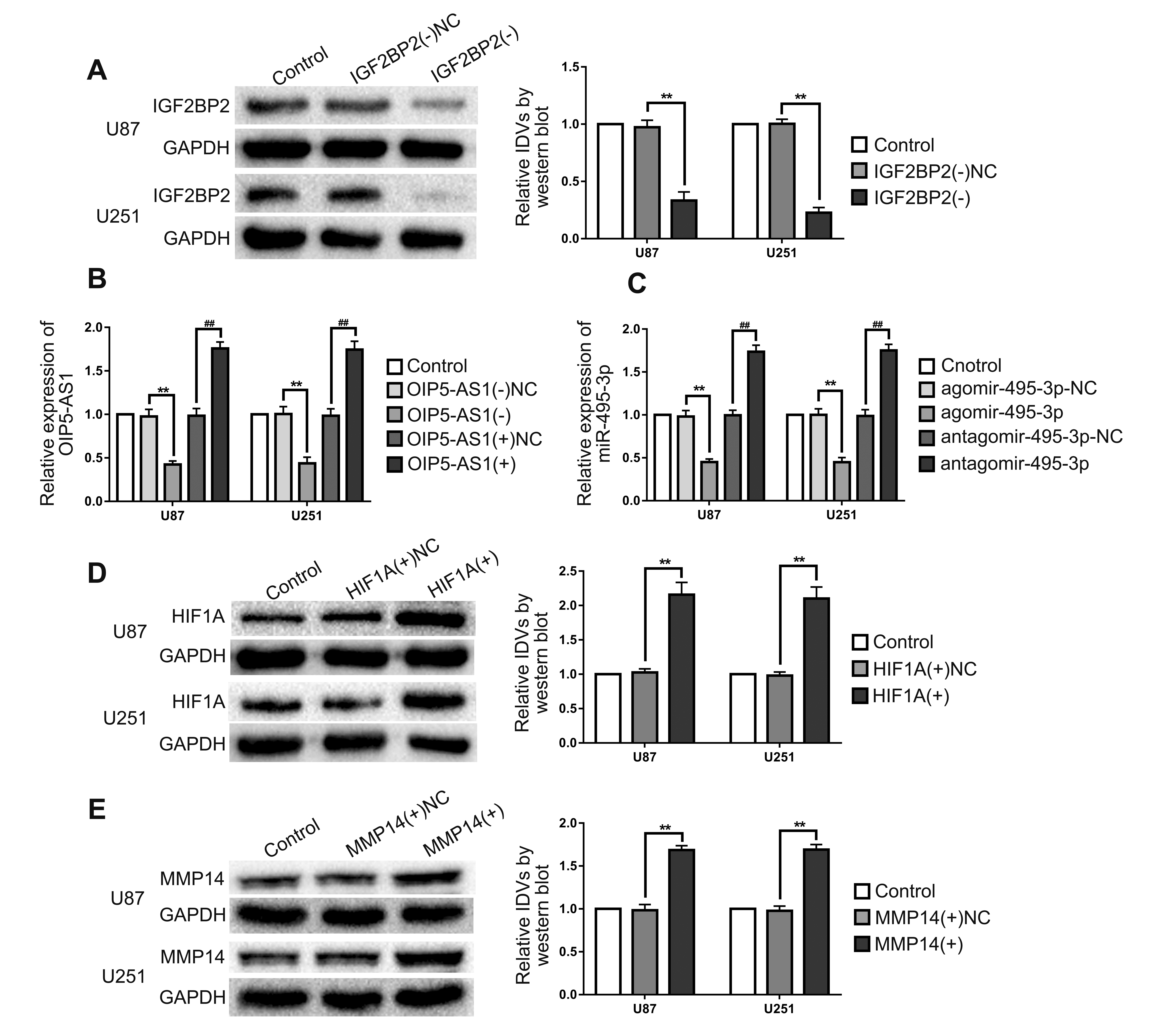
Western blot was used to detect the expression of IGF2BP2 in glioma tissues and cells. The results showed that in comparison to the NBTs, IGF2BP2 expression in LGGTs and HGGTs were significantly increased, and the expression increased as the pathological grade level increased; while IGF2BP2 expression in U87 and U251cellswere significantly higher than that in HA cells (Fig. 3A-B). To further explore the potential mechanisms of IGF2BP2 SUMOylation in regulating VM and the function of IGF2BP2 in glioma cells. The SUMO-site mutates (IGF2BP2-3KR) and knockdown IGF2BP2 cells were constructed. In comparison to the IGF2BP2-Wt group, cell proliferation, migration, invasion and VM of IGF2BP2-3KR group were significantly decreased and the expression of VM related proteins HIF1A and MMP14 also inhibited. Similar to above results, in comparison to the IGF2BP2(-)-NC group, cell proliferation, migration, invasion and VM of IGF2BP2(-) group were significantly decreased and the expression of VM related proteins HIF1A and MMP14 also inhibited (Fig. 3C-F).
OIP5-AS1 expression in glioma tissues and cells were detected by qRT-PCR. As the results showed, OIP5-AS1 was overexpressed in glioma tissues than in NBTs, and the expression increased with the pathological grade level. While the expression of OIP5-AS1 in U87 and U251 were significantly higher than that in HA cells (Fig. 4A-B). To further analyze the function of OIP5-AS1 in glioma cells, knockdown and overexpression plasmids ofOIP5-AS1 were transfected into U87 and U251 cells. In comparison to the OIP5-AS1(-)-NC group, cell proliferation, migration, invasion and VM of OIP5-AS1(-) group were significantly decreased and the expression of VM related proteins HIF1A and MMP14 also inhibited. On the contrary, cell proliferation, migration, invasion and VM of the OIP5-AS1(+) group were increased and the expression of VM related proteins HIF1A and MMP14 was also increased, by comparison to the OIP5-AS1(+)-NC group (Fig. 4C-F).
IGF2BP2 promoted VM in glioma cells by stabilizing and upregulating OIP5-AS1
This study further explored the effect of IGF2BP2 SUMOylated andIGF2BP2 knockdown on OIP5-AS1. The results revealed thatOIP5-AS1expression was significantly decreased by IGF2BP2SUMOylated andknockdown (Fig. 5A). In order to definite the relationship between IGF2BP2 and OIP5-AS1, RNA immunoprecipitation (RIP) assay was used to detect the direct binding between IGF2BP2 and OIP5-AS1. The results showed that OIP5-AS1 was significantly enriched in the anti-IGF2BP2 group by comparison to the anti-IgG group (Fig. 5B). Then, the regulation mechanism of IGF2BP2 knockdown on OIP5-AS1 was further elucidated by nascent RNA capture and mRNA stability assay. The results showed that IGF2BP2 knockdown did not affect nascent OIP5-AS1 but significantly reduced the half-life of OIP5-AS1 (Fig. 5C-D). Knockdown of IGF2BP2 and knockdown or overexpression of OIP5-AS1 plasmid co-transfection in U87 and U251 cells to further detect their functions in glioma.The results revealed that cell proliferation, migration, invasion, and VM of IGF2BP2(-)+OIP5-AS1(-) groupwere significantly decreased when compared with the IGF2BP2(-)+OIP5-AS1(-)-NC group.In comparison with the IGF2BP2(-)+OIP5-AS1(+)-NC group, in IGF2BP2(-)+OIP5-AS1(+) group, OIP5-AS1(+) reversed the inhibitory effect of IGF2BP2(–) on cell proliferation, migration, invasion, VM ability of glioma cells(Fig. 5E-G). Meantime, IGF2BP2 knockdown significantly inhibited HIF1A and MMP14 expression. Co-transfection of IGF2BP2 knockdown and OIP5-AS1knockdown enhanced the inhibition effect caused by IGF2BP2 knockdown alone, while co-transfection of IGF2BP2 knockdown and OIP5-AS1overexpression rescued the suppression effect (Fig. 5H).
MiR-495-3p were lowly expressed in both glioma tissues and cells, miR-495-3p agomir inhibited VM of glioma cells
QRT-PCR was used to detected miR-495-3p expression in glioma tissues and cells. The results confirmed that miR-495-3p expression was significantly down-regulated in glioma tissues and glioma cells (U87 and U251) (Fig. 6A-B). Up- or down-regulatemiR-495-3p to further explore the function of miR-495-3p in glioma cells. Cell proliferation, migration, invasion and VM of the agomir-495-3p group weresignificantly decreased, while the expression of VM related proteins HIF1A and MMP14 was inhibited. On the contrary, cell proliferation, migration, invasion and VM of antagomir-495-3pgroup were significantly increased, meanwhile, the expression of VM related proteins HIF1A and MMP14 was increased (Fig. 6C-F).
OIP5-AS1 targeted and negatively regulated miR-495-3p. Knockdown of OIP5-AS1 and overexpression of miR-495-3p inhibited VM of glioma cells
QRT-PCR was used to detect miR-495-3pexpression in glioma cells after OIP5-AS1 knocked-down. The results confirmed that miR-495-3p expressionlevel was significantly down-regulated in OIP5-AS1(-) group (Fig. 7A).The potential binding site between OIP5-AS1 and miR-495-3p was predicted by the bioinformatics database (Starbase 2.0).Dual-luciferase Reporter Gene Assays results indicated that the relative luciferase activity of OIP5-AS1-Wt in the agomir-495-3p group was significantly lower than in the agomir-495-3p-NC group, while in the agomir-495-3p-NC or agomir-495-3p groups, the relative luciferase activity of OIP5-AS1-Mut did not significantly change (Fig. 7B-C). These results indicate that OIP5-AS1 targets miR-495-3p. To figure out if miR-495-3p affected the tumor suppressor function of glioma cells induced by OIP5-AS1 knockdown, U87 and U251 cells co-transfected with OIP5-AS1(-) and agomir-495-3p or antagomir-495-3p. The results indicated that in comparison to OIP5-AS1(-) NC + agomir-495-3p NC group, cell proliferation, migration, invasion and VM of OIP5-AS1(-)+agomir-495-3p group were significantly decreased. The expression of VM-associated proteins HIF1A and MMP14 were significantly reduced (Fig. 7D-G).
MiR-495-3p inhibited the expression of HIF1A and MMP14 by directly binding their 3’UTR, thus inhibiting the VM of glioma cells
Up- or down-regulatedmiR-495-3p expression level in glioma cells to evaluated the the mRNA expression of HIF1A by qRT-PCR. The results indicated that HIF1A expression was significantly decreased aftermiR-495-3p agomir, while was significantly increased after miR-495-3p antagomir (Fig. 8A). The HIF1A 3’UTR existed a direct binding site ofmiR-495-3p which was predicted by the bioinformatics database (Starbase 2.0). Dual-luciferase Reporter Gene Assays results revealed that the relative luciferase activity of HIF1A-Wt in the agomir-495-3p group was significantly lower than in the agomir-495-3p-NC group, while in the agomir-495-3p-NC or agomir-495-3p groups, the relative luciferase activity of OIP5-AS1-Mut did not significantly change (Fig. 8B-C).To figure outif HIF1A could reverse the tumorsuppression effects of miR-495-3p on U87 and U251 cells, HIF1A (+) and agomir-495-3pwere transfected into cells and evaluated the biological behaviors. We found that the HIF1A overexpression significantly increased cell proliferation, migration, invasion and VM of glioma cells, and reverse the tumor-suppressive effect of miR-495-3p up-regulated (Fig. 8D-F). Similarly, similar results were observed in detecting the interaction between MMP14 and miR-495-3p (Fig. 8G-L).
The inhibition of SUMOylation of IGF2BP2 could inhibit tumor xenograftgrowth, prolong the survival time and yield the lowest VM formationof nude mice
The function of IGF2BP2 SUMOylation on glioma cells was further determined by using the subcutaneous and orthotopic transplantation model. As the results presented, in comparisonto IGF2BP2-Wt group, the tumor volumes were significantly lower in IGF2BP2-3KR group (Fig. 9A-B).In addition, survival analysis showed that nude mice in IGF2BP2-Wt group have a longer survival time than in the IGF2BP2-3KR group (Fig. 9C). Last, the CD34-PAS dual-staining was used to detect the VM in different groups. The VM density in IGF2BP2-3KR group was lower than that in IGF2BP2-Wt group (Fig. 9D).
{kind=link}