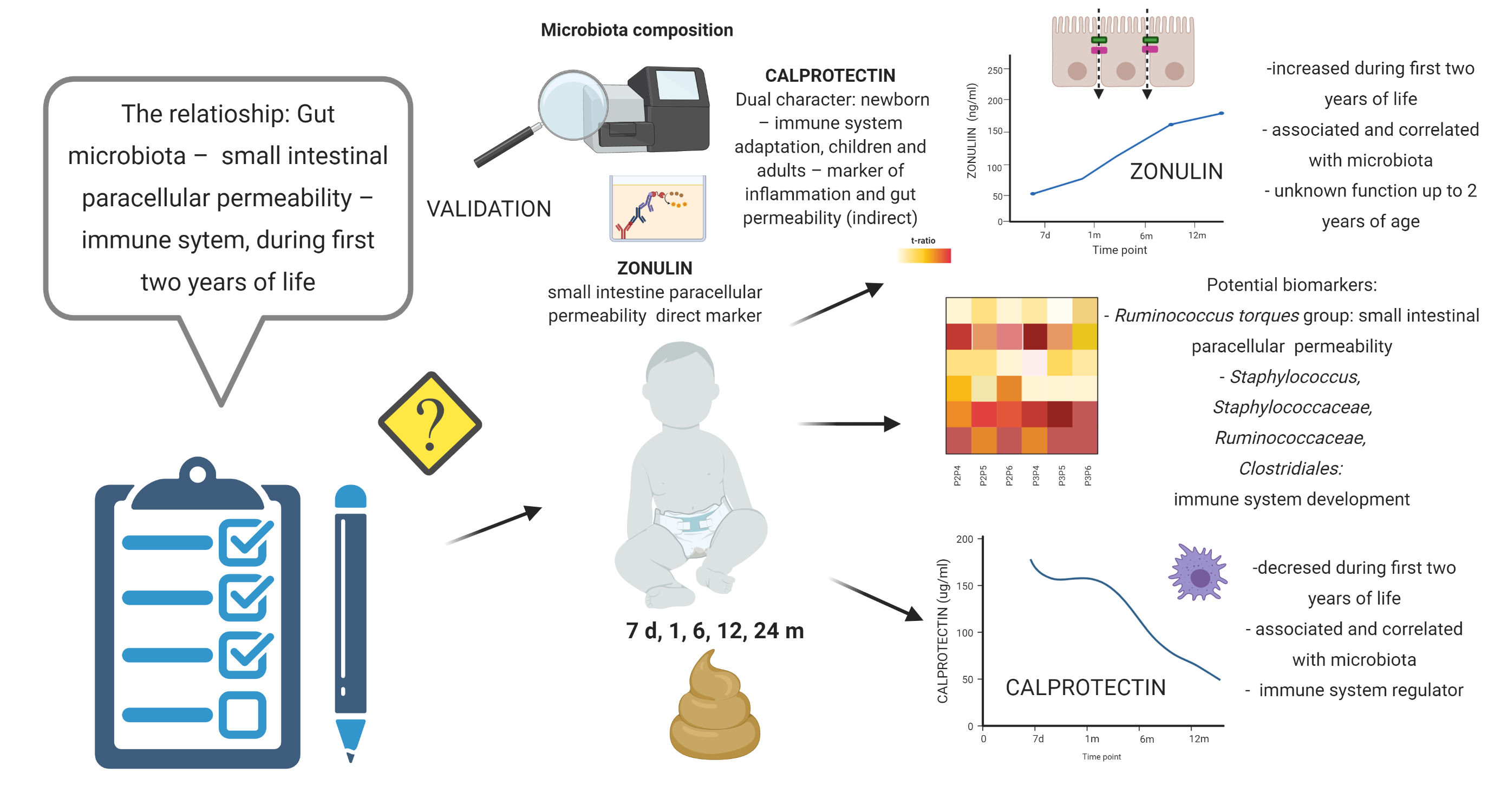
This is the first study in which putative associations between the gut microbiota and the concentrations of zonulin and calprotectin in children's stool during the first two years of life have been investigated. The study of zonulin allows the non-invasive assessment of the functional state of the small intestinal paracellular permeability; additionally, the calprotectin measurements provide insight into the development of the immune system after birth. The observed results indicate that small intestinal paracellular permeability measured based on zonulin concentration is higher in the period from the sixth month to the second year of life than in the first months of life. In line with this, the concentration of calprotectin in stool in the first and second years of life is lower than that in the first months of life; of note, the concentration of calprotectin at six months of age does not differ significantly from that in the previous or later adjacent time points. Since the validation data also revealed a difference in the concentration of calprotectin between the first and sixth months of life, this allows us to define the 6th month of life as the key point for forming the small intestinal barrier and for the development of postnatal immunity. It should also be stressed that, usually, in the sixth month of life, solid food is introduced to children’s diet, which can significantly impact both gut microbiota and intestinal permeability.
Although zonulin seems to be a valuable marker of small intestinal paracellular permeability, the data in the context of children aged two years old or younger are scarce. Tarko et al.[37] reported significantly higher blood zonulin concentrations in children with gut wall defects and infected with rotaviruses. Saleem et al.[38] observed that the blood zonulin levels were significantly lower in children delivered before 28 weeks of pregnancy. The fecal zonulin concentration in children during first two years of life was not reported and its role during this period is unknown. Its relationship with gut microbiota seems to be interesting.[38] Zonulin is involved in controlling the passage of molecules weighing at least 3.5 kDa [39] through the intestinal barrier via its reverse influence on TJ tightness.[40, 41] After the activation of zonula occludens (ZO) 1, zonulin controls the low-capacity “leak” type route characterized by low selectivity.[42, 43] Interactions between zonulin and gut microbiota can be bidirectional. Bacterial and gluten exposure was defined as intensive triggers of zonulin release.[19, 44] Of note, the zonulin pathway is an innate defensive mechanism of the host, able to control the gut microbiome composition via the “flushing out” of microorganisms by water secreted into the intestinal lumen following hydrostatic pressure gradients.[45] High stool zonulin levels suggest that the gut barrier allows the free flow exchange of various particles in infants. Moreover, it is likely that the commercially available ELISAs detect one or more members of the zonulin family that have not been discovered yet but play some important role during the first two years of life.[46]
High levels of calprotectin in feces during the first months of life are associated with the regulation of the development of the immune system in neonates as well as with the adaptation to new environmental conditions.[25] The decrease in the fecal calprotectin concentration implies that inflammatory processes in the gut tend to decrease from the sixth month of age, which, with the accompanying increased paracellular intestinal permeability and bacterial alpha diversity, provides evidence of immune tolerance. Willers et al. [25] demonstrated that mice exposed to calprotectin immediately after birth induced microbial tolerance against the first wave of microbial colonization; on the other hand, the same exposure after the neonatal period was associated with pro-inflammatory responses. Our results did not fully corroborate these observations. The intestinal permeability in the first month of life, as per the zonulin concentration, is lower than that in the later period, suggesting that the possibility of antigen translocation early after birth is limited. After the sixth month of age, the intestinal paracellular permeability increases, but this does not increase inflammation, as per the calprotectin concentration, which indicates the development of immune tolerance. The differences between the two studies may be due to the fact that Willers et al. did not directly measure intestinal permeability and additionally used an animal model that does not closely mimic the timing of human intestinal barrier development. Moreover the role of paracellular way in translocation of bacteria and endotoxin is still the subject of debate[22], what requires that the observed results should be interpreted with caution.
In summary, the increased production of zonulin may be caused by changes in intestinal bacteria and by the introduction of gliadin into the diet. In contrast, the decrease in the calprotectin content in the stool may indicate immunological tolerance development. However, it should be remembered that breast milk is an essential source of calprotectin, which can significantly influence the fecal calprotectin content. This said the observed changes cannot be explained solely by environmental factors as zonulin decreases in the later years of life despite the constant consumption of gliadin. Therefore, in the context of longitudinal analyses of biomarker trajectories, we applied corrections for the time of breastfeeding and the type of delivery to minimize the influence of these confounders. To help resolve the mechanisms underlying our results, we decided to analyze whether a cause-effect relationship between the microbiota and the biomarkers would be observed. Of note, the microbial changes in the first two years of life are very dynamic and depend on the mode of delivery, feeding, antibiotic treatment, and other environmental factors.[28, 29] However, a comprehensive analysis of all factors influencing the gut microbiota is beyond the scope of this study, which is focused on the relationship between the microbiota and the fecal zonulin and calprotectin content. This said the compositional and functional comparison of the PMU and HMS cohorts' microbiota would be very interesting.
The Shannon index (more sensitive to species richness), shared 5 and 4 significant time intervals with zonulin (increasing) and calprotectin (decreasing), respectively. Similar results were observed for coordinates of the first two axes in the context of a principal coordinate analysis based on the Bray-Curtis distance, one of beta-diversity measures. However, it translated only into a negative correlation between alpha-diversity indices, the Shannon index in particular, and calprotectin. This negative correlation may be associated with the development of immunological tolerance. The microbial colonization of the intestine after birth will generate a diversity of new antigens that will play an important role in the stimulating of epithelial function and the establishing the offspring’s immune system.[47] Of note, weaning (the average breastfeeding time in our study was about 30 weeks), and the resulting loss of milk-borne calprotectin, sIgA and other anti-microbial factors, along with the transition from mothers’ milk to a complex diet (including solid food), can have a major impact on the dynamic microbiota development, and thus in the resulting immune responses during the neonatal period.[6]
Willers et al. [25] reported that the gut microbiota's overall diversity significantly increases during the first year of life, similarly to what we observed in this study. Moreover, they observed the increased abundance of the bacterial classes Actinobacteria, Bacteroidia [48–50] and Clostridia, along with the decreased abundance of Bacilli and Gammaproteobacteria over the same time period. Of note, Willers et al.[25] demonstrated that during infancy, high abundance of Actinobacteria and low abundance of Gammaproteobacteria are linked to high fecal calprotectin levels; additionally, at the family level, fecal calprotectin promoted the higher abundance of Bifidobacteriaceae and the reduction of Enterobacteriaceae via the production of acetate[51]. Taxonomic composition observed by Willers et al.[25] might be linked to the elevation in the synthesis and reduction in the degradation of short-chain fatty acids (SCFAs) and therefore leading to the overrepresentation of health-promoting gut microbiota metabolic functions. Our results confirm the observations obtained by Willers et al.[25] in terms of abundance; however, the correlations in the cited study were not reproduced here. Importantly, the observed differences may depend on the difference between the investigated populations (Polish and German) or/and the analytical methods used. This said the analysis of both data-sets showed similarities with respect to the correlation between the levels of calprotectin and bacteria at the family level.
Interestingly, the bacteria associated with zonulin and calprotectin (Supplementary Table 5) may be involved in the production of SCFA, which is a protective factor in the context of intestinal barrier integrity and of inflammatory processes. Particularly, the different taxonomic groups of bacteria within the class Clostridia were associated with increased zonulin and decreased calprotectin concentrations. Within this class, the family Ruminococcaceae was more abundant in women with low zonulin concentrations in a previous study; however, here, we observed the opposite pattern. Importantly, not all bacteria within this class have the same properties. For instance, the genus Ruminococcus (gnavus group) – positively correlated with fecal zonulin concentration, is increased in patients with IBS-D and Crohn’s disease, which suggests its pro-inflammatory activity. An interesting result was obtained in the case of the Ruminococcus (torques group). Ruminococcus torques, a butyrate producer, have been found to dominate patients' gut milieu with Crohn’s Disease.[52, 53] They were also studied in the context of individuals with increased risk of upper gastrointestinal tract involvement, but with inconclusive results.[54–56] Importantly, Ruminococcus torques are able to utilize mucin in the human intestine, sustaining their adaptability to the human intestinal environment.[53, 57, 58] Consequently, increased mucin degradation makes luminal antigens cross the gut barrier and stimulate the immune system, leading to intestinal disorders.[59] Of note, in this study, the abundance of these above bacteria positively correlated with the concentration zonulin and the dynamics of calprotectin concentration. Therefore, bacteria from the Ruminococcus (torques group) may include species associated with small intestinal permeability and immune responses in the first two years of life. For deeper species resolution and assessment of the gut functional potential future shotgun metagenomics, metabolomics, and immune analyses will be essential.
In this study, the Bacilli class correlated with decreased zonulin and increased calprotectin levels. In general, this class is considered beneficial for gut health, which is in line with decreased zonulin levels. On the other hand, the family Staphylococcaceae and genus Staphylococcus from the Bacilli class are the most prominent bacteria in human milk, an important calprotectin source in the infant. Family Enterococcaceae belongs to opportunistic pathogens, considered as potential marker for IBD what can explain correlation with increased fecal calprotectin concentration. However, our present results do not allow consistent conclusions on the role of bacteria in the small intestinal barrier's permeability or the immune system in the first two years of life. For example, the Lachnoclostridium genus produces SCFAs and occurs in lower amounts in patients with gastrointestinal cancers; therefore, in theory, it should decrease intestinal permeability; however, it was correlated with increased concentrations of zonulin in the neonatal stool in this study. Alphaproteobacteria occur in high numbers in patients with IBD and are negatively associated with the concentration of zonulin. The order Corynebacteriales is associated with decreased zonulin and belongs to the class Actinobacteria, which was positively correlated with calprotectin in the study by Willers et al.[25] The order Pseudomonadales and the family Moraxellaceae are negatively correlated with zonulin and belong to Gammaproteobacteria, negatively correlated with calprotectin in the study by Willers et al. Therefore, in no case was there a full consistency of microbiota changes between particular time points and changes in the levels of zonulin and/or calprotectin. This said the validation analysis suggested that the genus Staphylococcus and the family Staphylococcaceae (positively correlated with the concentration of calprotectin), as well as the family Ruminococcaceae and the order Clostridiales (negatively correlated with the concentration of calprotectin), are potential candidates as markers of the development of the immune system in children under two years of age.
Finally, we analyzed the metabolic pathways linked with bacterial abundance (Supplementary Table 6). We were able to show that the pathways involved in the production of SCFAs and the metabolism of carbohydrates were positively associated with fecal zonulin and negatively with calprotectin. The latter association suggests the role of SCFA in the immune response. However, the positive associations and correlations between pathways involved in SCFA production and the zonulin elevation are difficult to explain, suggesting that other mechanisms are also involved in regulating small intestinal barrier permeability in children of up to two years old. It should be emphasized that the variations in the concentrations of zonulin and calprotectin overlapped with the introduction of new foods to the child’s diet around the sixth month of life (addition of soup, grated apple), with the consequent alteration in the gut microbiota composition and the related metabolic functions. For example, metabolic pathways involved in the metabolism of carbohydrates, vitamin B6 and nucleotides positively correlated with the levels of zonulin, which could suggest that an “open” small intestinal paracellular permeability could play a role in the transportation of metabolic reaction products during intensive growth and development. However, the paracellular transport of nutrients is known to play a rather minor role. The observed results support the concept that paracellular permeability is only the component of intestinal barrier[22] and that SCFAs affect mostly transcellular route via metabolism of enterocytes. Of note, the analysis of the validation data did not confirm our observations concerning the metabolic pathways. Therefore, shotgun sequencing and metabolomic analyses should be performed to shed more light on these processes.
Our study is not without limitations. Our cohort was relatively small, though enough to observe changes in zonulin and calprotectin levels as well as in gut microbiota composition during the first two years of life with appropriate power. Of note, our results were validated using the HMS cohort as well as via the comparison between the microbiota of meconium and stool samples collected at other time points. Moreover, our samples were somewhat heterogeneous (with respect to the delivery mode, the use of antibiotics, nutrition, and other environmental factors). Of note, all of these factors can affect the gut microbiota; however, in term infants, the mode of delivery seems to impact fecal calprotectin levels only in the first week of life.[25] Moreover, in our study, we did not use direct methods to measure the gut permeability; we used biomarkers, for which data in healthy children < two years old are limited. In fact, there are studies reported on the fecal levels of zonulin in adults but not in children. It is known that in children, the levels of zonulin and calprotectin are incomparably higher than those in adults, changing over time, and could be influenced by e.g., breastfeeding, the dietary patterns, the mode of delivery, and the consumption of gliadin. In the current study, however, due to the small sample size, we did not analyze these biomarkers separately in the context of different delivery modes. Also, we did not collect detailed data on antibiotic exposure. Further, we did not measure these factors in blood (ethical reason - healthy children); only in stool. Moreover, since no metabolomic, immunological, or shotgun metagenomic analyses were performed, the results we obtained must be treated with caution. Mechanistic studies are still needed to investigate the relationship between microbiota and gut permeability and the immune system.
{kind=link}