Following the guidance from major regulatory agencies, which requires identification, safety, and efficacy assays [1, 40, 41], the present work completed a series of tests to assess the probiotic potential of nine novel Brazilian L. plantarum strains. All strains had their identification confirmed by MALDI-TOF, and this finding was supported by classical microbiological identification (data not shown).
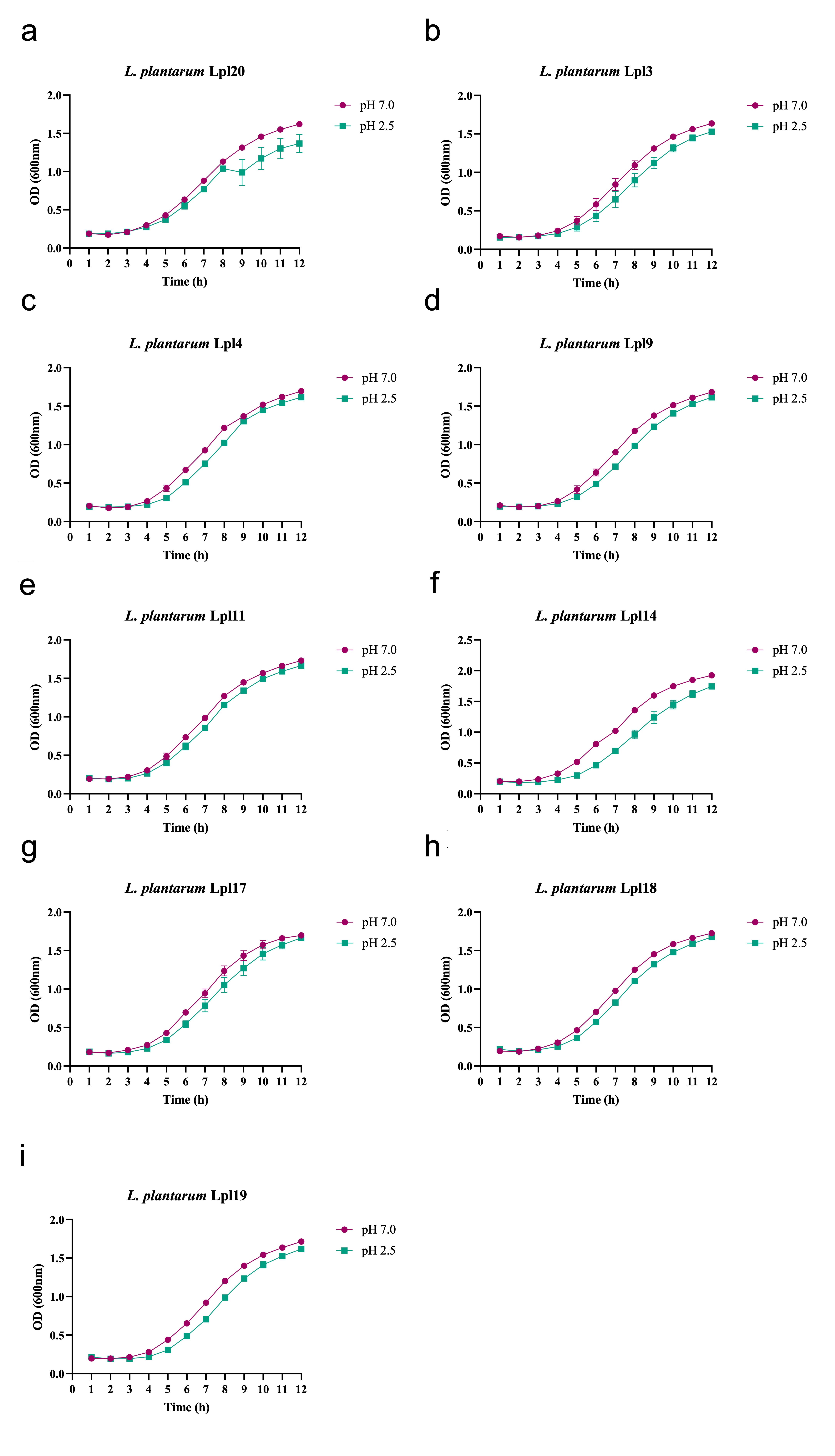
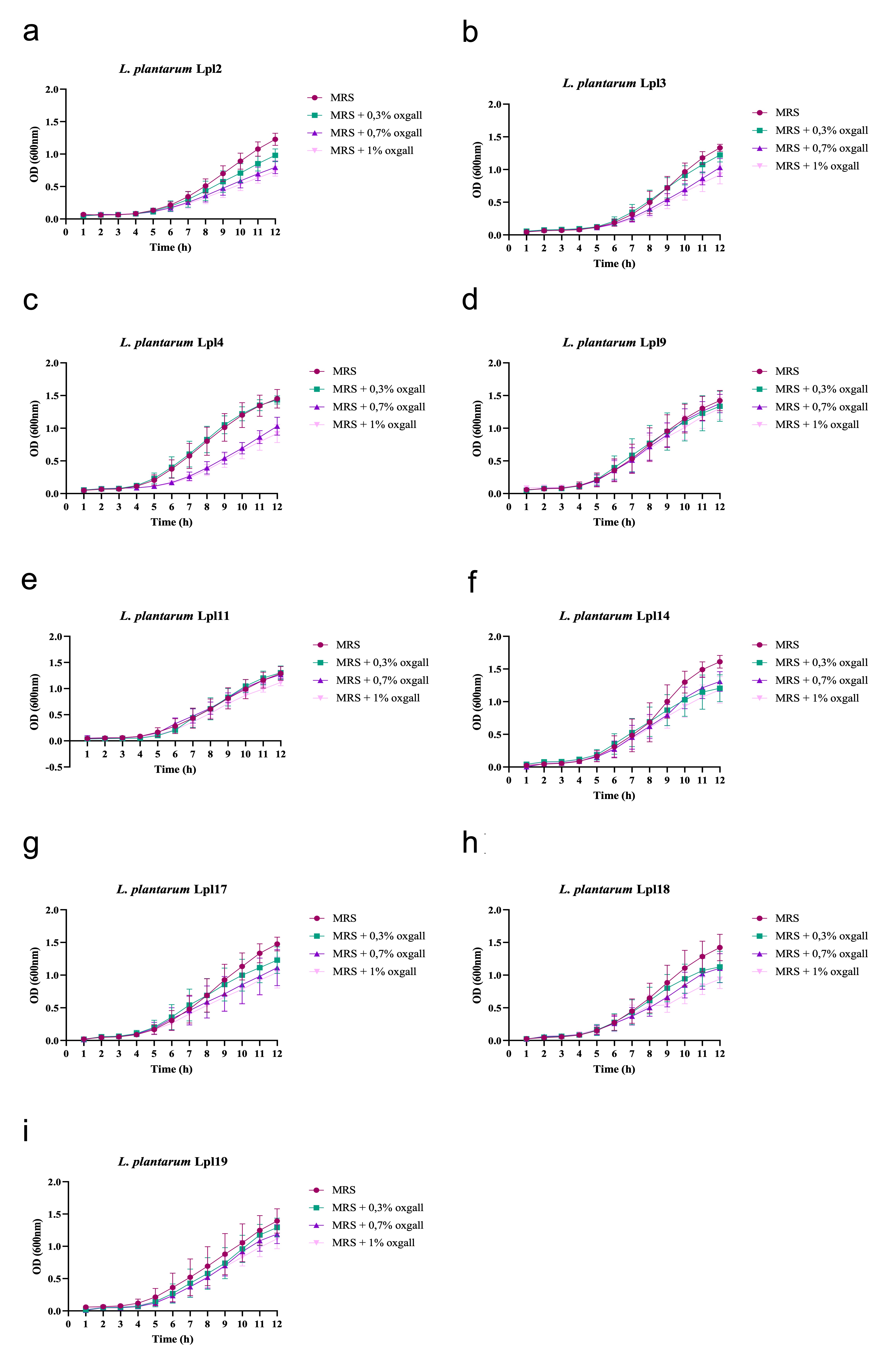
After having their identification confirmed, it is recommended that novel probiotic candidates be able to tolerate acidity and bile salts during transit through the GIT [42]. In this work, L. plantarum strains showed increased tolerance to acidic pH (> 80 %)and bile salts (> 70 %) thus signalling that they might be able to reach the intestine in great number to promote their beneficial effects. These findings are supported by previous studies which showed that strains within this genus are generally tolerant to harsh GIT conditions. Haghshenas and collaborators [31], revealed that the resistance of L. plantarum 15HN after 3 hours of incubation at pH 2.5 is around 71 to 76% [31]. More recently, Nath et al. [43], demonstrated that L. plantarum strain GCC_19M1 could survive at rates of 93.48–96.97% when submitted to pH 3.0 for 3h [43]. Similarly, L. plantarum 15 was able to tolerate gastric pH (pH 2.0), with a survival rate of 86.40% [44]. Regarding bile salts tolerance, Jacobsen et al.[45] compared the resistance of 47 Lactobacillus strains, including L. plantarum, L. rhamnosus, L. acidophilus, and L. crispatus, and observed that most of them had relatively high resistance to bile salts when incubated for 4h in 0.3% oxgall [45]. Similar results were found for L. plantarum H3b and K3b[46] and L. plantarum strain GCC_19M1 [43], which were able to survive in similar ratios when exposed to 0.3% of bile salts.
Previous studies have demonstrated that some LAB can inhibit pathogens' growth, such as Shiguella sonnei [47], Listeria monocytogenes [48], E. faecalis pathogenic [49] and Escherichia coli [50], which can be a promising probiotic feature. In this study, it was observed that Brazilian L. plantarum strains have a broad inhibition spectrum against commonly gastrointestinal pathogens. These findings are supported by other studies, which observed that other L. plantarum strains were also able to inhibit potentially pathogenic bacteria, such as E. coli, Staphylococcus aureus, E. faecalis, L. monocytogenes [51], Bacillus cereus, Acinetobacter johnsonii, and Pseudomonas aeruginosa strains [43]. This inhibition may be due to the production of H2O2, organics acid production as lactic acid or acetate, and bacteriocins as plantaricin [52–54]. Such mechanism supports their application as food preservatives or as a potential alternative antimicrobial strategy in the actual context of increasing antibiotic resistance among pathogens. [55–57]
Probiotic bacteria candidates should also possess the ability to adhere to the intestinal mucosa, which could potentially result in transient colonisation, enhancing immunomodulatory effects and inhibiting pathogen growth through the mechanisms of competitive exclusion and bacterial antagonism [58–60]. In the present study, it was observed a high adhesion rate to Caco-2 cells by Lpl2, Lp9, Lp11, Lpl17, and Lpl19. Sharma et al. [61] reported Caco-2 cell adhesion values ranging from 2.45 to 9.55% when studying lactic acid bacteria isolated from fermented foods [61]. Garcia-Ruiz et al. [62] also analysed probiotic bacterial strains isolated from wine and obtained adhesion levels to Caco-2 cells ranging from 0.37–12.2% [62]. Behbahni et al. [63], reported adhesion values of 12.2% when analysing the adhesion of L. plantarum L15 to Caco-2 cells [63]. Those studies corroborate the results obtained here, as the variation between adherence rates was between 0.72–13.64 %. The potential to adhre and interact with epithelial cells by probiotic bacteria allows them to regulate the activation of the host’s immune system. For example, Taverniti et al. [64] demonstrated that L. helveticus MIMLh5 reduced the activation of NF-κB in Caco-2 cells [64]. Other studies showed that pro-inflammatory cytokines Il1b and TNF, and anti-inflammatory cytokine TGFβ were induced by L. sakei, and L. johnsonii, respectively [65].
Safety evaluation of new probiotic candidates for human and animal applications should also be performed. Thus, in this study, we have evaluated the antibiotic resistance profile, mucin, and haemolytic activity of L. plantarum strains. None of them presented haemolytic or mucolytic activity, demonstrating the absence of potential risk for the host epithelial barrier disruption or haemolysis [33, 66, 67]. These findings are supported by other studies that demonstrated no haemolytic or mucolytic activity by LAB potential probiotic strains such as L. delbrueckii CIDCA 133 [68], L. plantarum BCC9546 [69] and L. helveticus D75 [70].
Regarding antibiotic resistance, some L. plantarum strains presented a resistance profile to gentamicin, streptomycin, and vancomycin. Lactobacillus species are known to be intrinsically resistant to vancomycin, as observed in all strains in this work, but this constitutive resistance is not considered a matter of concern [71–73]. Corroborating with the findings in the present study, Hummel et al. [74] found that among forty-five LAB strains (including Lactobacillus genera) aminoglycosides (gentamicin and streptomycin) and ciprofloxacin resistance was prevalent in more than 70% of them, indicating that maybe this could be an intrinsic resistance [74]. In the same study, a low resistance profile was observed for ampicillin, penicillin, chloramphenicol, and tetracycline [74]. Anisimova et al. [71] analysed the antibiotic resistance profile of twenty Lactobacillus strains and showed that 90% of them were sensible to clindamycin and 95% sensible to tetracycline, erythromycin, and rifampicin. All of them were inhibited by chloramphenicol and ampicillin. As expected, the majority were resistant to vancomycin and aminoglycosides [71]. It should be highlighted that antibiotic resistance becomes a problem when resistance is transferable by mobile elements such as plasmids [40, 72, 73]. This shows that besides phenotypic profile, genotypic methods should also be applied to study the whole antibiotic profile due to potential intrinsic or transferable resistance genes [75], thus being one of the future goals of the present work.
After passing through in vitro characterization, we also accessed the efficacy of the present strains in vivo, through an experimental mice model of 5-FU (300mg/kg)-induced intestinal mucositis. This well-established model was chosen due to its severity, which would make it possible to observe more significant protective effects promoted by potential probiotic strains [76, 77]. Additionally, it is important to highlight that those studies evaluating the effect of L. plantarum in the mucositis model are lacking, with the main findings provided by Levit et al. [27] with L. plantarum CLR2130, and Ciobanu et al. [28] with L. plantarum ATCC 8014 [27, 28]. Therefore, this work provides important information about how different L. plantarum isolates can exert a beneficial impact in this model.
It was observed mice body weight loss, epithelial architecture alteration, villus shortening, and polymorphonuclear cell infiltration in the intestinal mucosa after administration of 5-FU (300 mg/kg). These findings are consistent with the pathobiology of mucositis induced by chemotherapy [22, 35, 76, 77]. Consumption of all Brazilian L. plantarum strains were able to reduce eosinophils peroxidase and myeloperoxidase activity, demonstrating their beneficial effect to protect the intestinal mucosa from polymorphonuclear cells infiltration. Additionally, most strains were able to protect against villous atrophy and improved histopathological score (Lp12, Lp13, Lp14, Lp11, Lp19 and Lp18). None of the strains were able to prevent weight loss. These results are supported by other studies which observed improvements in some mucositis parameters after probiotics administration regardless of improvement in water/feed consumption or bodyweight [18–20, 37].
Soares and Sonis et al.[36, 77] suggested that up-regulation of pro-inflammatory cytokines including Tnfα, Ifnγ, and Il6 might be involved in mucosal injury severity in acute stages [36], with Tnfα also related to the amplification of inflammatory response in 5-FU-induced mucositis [77]. In this study, we demonstrated that administration of some L. plantarum promoted down-regulation of proinflammatory cytokines Il6 and Il1b (Lpl4, Lpl9, Lpl11, Lpl14, Lpl17, Lpl18 and Lpl19). Some strains were also able to up-regulate gene expression of the immunoregulatory cytokine Il10 (Lp12, Lp19, Lp14, Lp17 and Lp18), a potential anti-inflammatory cytokine related to being involved in the prevention of damage during intestinal inflammatory processes [78, 79]. These findings are corroborated by previous works that showed that administration of probiotics such as L. acidophilus [80, 81], L. casei [18], Bifidobacterium sp. [20], L. plantarum strains[27, 28] were able to decrease inflammatory cell infiltrate and improve the inflammatory and functional aspects of intestinal inflammation induced by 5-FU through expression modulation of inflammatory cytokines Il6, Ifnγ, Il1b, Tnf, and Il10. Thus, the result of the present study further highlights the anti-inflammatory effect that Brazilian L. plantarum strains can exert on the host, ratifying their beneficial effects and use as probiotics. In addition, besides having a positive impact on the improvement of mucositis in the murine model, some studies have demonstrated that the association of two or more probiotic strains can enhance their effects [20, 82, 83]. Therefore, the strains characterized in this study can also be associated to maximize their effects, opening a wide range of studies and possibilities to be explored.
In conclusion, the present study revealed the strain-dependent functionality and host beneficial effects of L. plantarum strains isolated from different sources in Brazil. Most strains tested in vitro appear to be safe and presented good features, regarding probiotic potential, to be explored in different fields, such as in the food industry, biotechnology, and nutrition. Additionally, most of them were able, at some points, to alleviate mucositis symptoms in the mouse model. This study opens a wide range of possibilities to be explored as it provides evidence to support further studies for the application of novel L. plantarum strains.
{kind=link}
{kind=link}