The treatment of RGM infection is often very difficult because of their higher drug resistance rate than SGM and unavailability of highly potent drugs against them in vitro. M. abscessus complex and M. fortuitum are two most prevalent RGM species around the world. Infections due to M. abscessus carry a poor prognosis since this RGM is, for all the correct reasons, considered an “antibiotic nightmare”(16). Thus, identifying drugs that could work potently against M. abscessus is a priority. M. massiliense is a species that originally splits from M. abscessus but it is located closely in the phylogenetic tree (17). The treatment response rates to clarithromycin-based antibiotic therapy are much higher in patients with M. massiliense than patients with M. abscessus lung disease (18). M. fortuitum is the main RGM responsible for extra-pulmonary disease, especially in cutaneous and plastic surgery-related infections(19). In contrast to M. abscessus, M. fortuitum infection has better prognosis due to some available effective drugs(20). However, its emerging drug resistance highlights the need for new and effective drugs(20–22). Several studies have verified the efficacy of LZD in MDR-TB or even in XDR-TB treatment (9, 13, 23). A few studies also proved its antibacterial activity against NTM species either in vitro or in vivo (24, 25). As a novel oxazolidinone prodrug, TZD exhibited greater potency than LZD against M. tuberculosis (6, 26) as well as against NTM(27, 28). Limited studies or no study has been performed to evaluate the efficacy of SZD and DZD against NTM species (28), whereas only a few studies provided preliminarily assessment of their potential usage in TB (14, 29, 30). In this study, we evaluated the efficacies of four oxazolidinones against the reference and clinical isolates of RGM to gain insights into their potential use for specific RGM species.
As new drugs, well recognized susceptibility testing methods for TZD, SZD and DTD have not been developed and the breakpoints to define drug resistance for them have never been discussed yet. Therefore, the MIC data of different RGM species against oxazolidinones still remain scarce. In this study, the four oxazolidinones exhibited promising activities in vitro against the recruited RGM reference stains. The absolute majority of species had MICs below 8 µg/mL against the four drugs. However, different species presented non-uniform susceptibility patterns. The MIC distributions of M. massiliense had similar tendency to the M. abscessus, but the MICs of TZD were obviously higher than M. abscessus. Brown-Elliott et al demonstrated M. abscessus is more resistant to TZD, compared with M. massiliense with 2-fold lower MIC90 and MIC50 (28).However, Ruiz et al showed the MIC50 of M. massiliense against tedizolid was two fold higher than that of M. abscessus (6).The main reason for the this discrepancy is sampling error, the constituent ratio of M. abscessus and M. massiliense was different among studies.Furthmore, the resistant profile of stains isolate from different countries may be not exactly the same.In comparison with other oxazolidinones, the MIC values for TZD were the lowest for both M. abscessus and M. massiliense. Previous studies, including 170 isolates of RGM, showed equivalent or lower (1 to 8 fold) MIC50 and MIC90 values for TZD in contrast with LZD(28). Furthermore, TZD harbors several advantages over LZD in terms of tolerability, safety, dosing frequency, and treatment duration(31). Only a few studies have reported the clinical use of TZD for the treatment of NTM infections. Our results indicated that its usage seems reasonable for the treatment of infection caused by M. abscessus and M. massiliense. Among the 25 tested M. fortuitum isolates in our study, 22 (22/25) strains had MICs of LZD at ≥ 16 mg/L. Based on the CLSI criteria, these strains could be categorized as intermediate resistant or resistant strains, 52% (13/25) of them belong to resistant strains. Using the cutoff value of LZD as the tentative breakpoints, the susceptibility rate of M. fortuitum against TZD, SZD and DZD were 20% (5/25), 12% (3/25), 76% (19/25), respectively. DZD exhibited the best antimicrobial activity against the M. fortuitum. However, whether this in vitro outcome reflects the in vivo efficacy or not, requires further investigation.
A major limitation of this study was that no recommended breakpoint of different NTM species against TZD, SZD or DZD had been proposed previously. Besides in vitro MIC distributions, the breakpoint determination also correlates with clinical treatment response and pharmacokinetic/pharmacodynamics (PK/PD) data including drug dose. The clinical trials on these new oxazolidinones are either unavailable or very limited. A few studies have been performed on the pharmacokinetic analysis of these drugs. Generally, all the drugs were well-tolerated, and the Cmax were highly dose-dependent. Recently, Choi et al demonstrated that, after multiple doses of TZD up to 1200mg twice daily for 21days,the peak serum concentration was 16.3 µg/mL, which is comparable with peak serum concentrations of LZD = 12.5 µg/mL at the dosage of 300mg twice daily(15, 32). In another study, a single 800mg dose of DZD under fasting condition acquired Cmax at 11.74 µg/mL(33). STD presented superior efficacy than LZD against experimental murine model of tuberculosis. The Cmax of its major metabolite PNU-101603, which contributes to its activity, was 6.46 µg/mL at given a dose of 1200mg QD (40). However, since the optimal dosage of these next-generation oxazolidinones is still under investigation, the appropriate breakpoints for the susceptibility definition of these drugs remain beyond known.
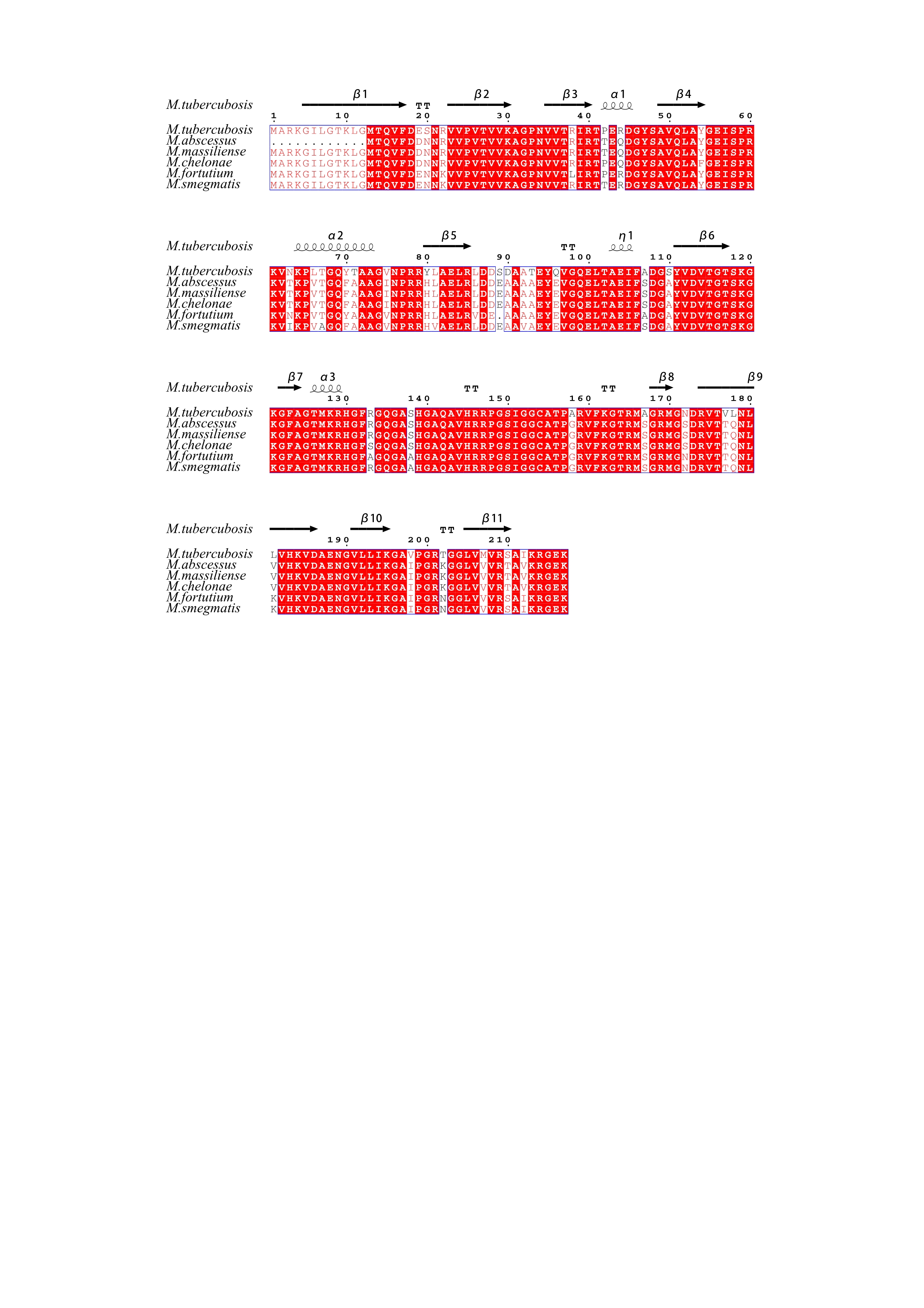
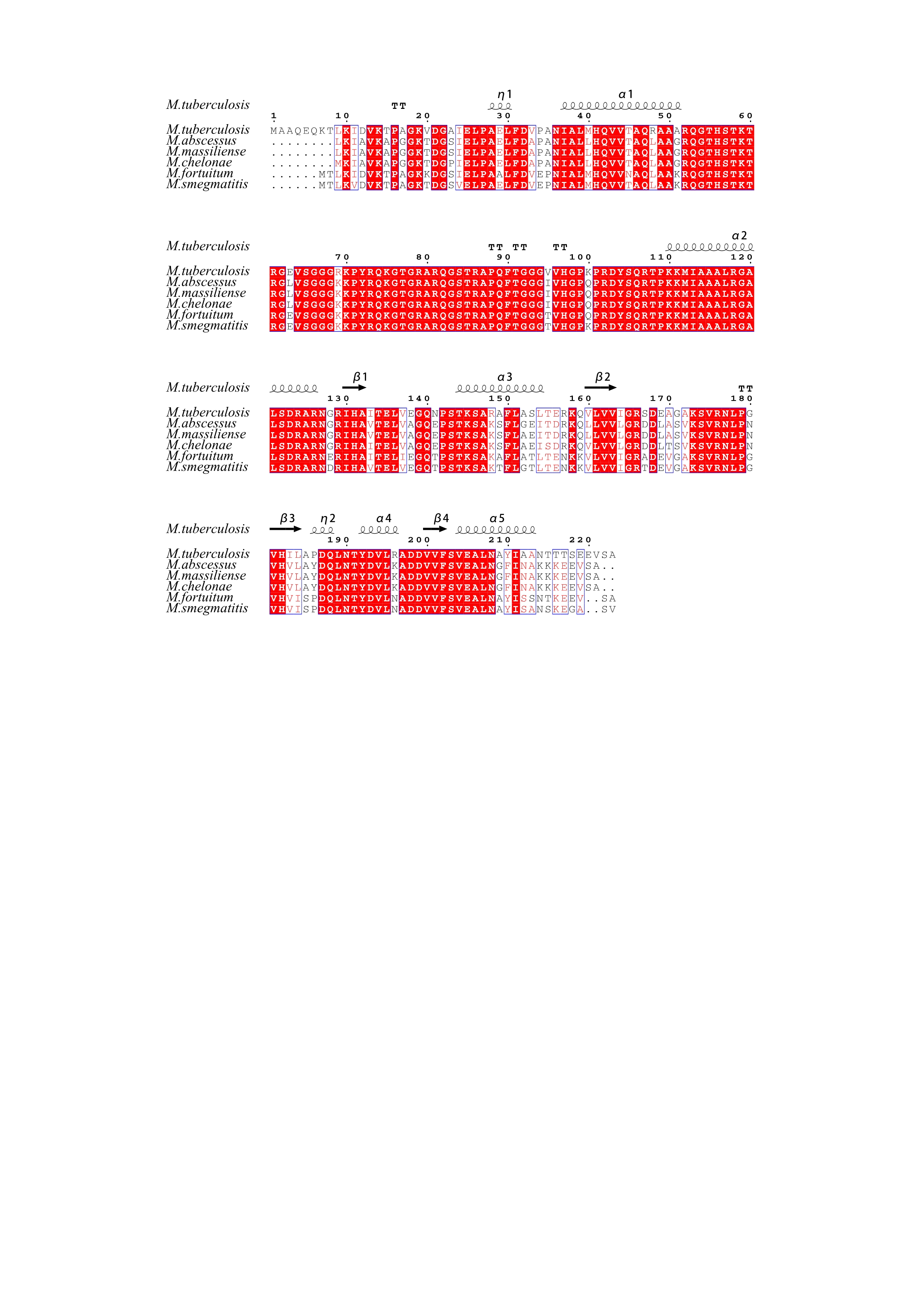
LZD works by binding to the peptidyl transferase center of the 50S ribosomal subunit, which is composed of 5S and 23S rRNAs and 36 riboproteins (L1 through L36)(34). Recently, the Cryo-EM structure of the large ribosomal subunit from M. tuberculosis bound with a potent LZD analog (LZD-114) was determined(35). LZD-114 is similar to LZD in C ring but different in A and B ring, in that it lacks a fluorine group in the B-ring while the original morpholine ring is replaced by a thiazole ring in the A-ring (Fig. 3A). The LZD-114 is also bound in the same pocket and in a similar orientation to LZD in other species (36, 37). The structure showed that rplC encoded ribosomal protein L3 and rplD encoded ribosomal protein L4 bound directly to 23S ribosomal RNA and was placed relatively close to the LZD binding site on the ribosomes, suggesting that the mechanism for reduced susceptibility may include structural perturbation of the LZD binding site (PDB:5V7Q). Furthermore, previous studies demonstrated that mutations in rplC and rplD could lead to LZD resistance in M. tuberculosis(12, 38). However, there is no non-synonymous mutation in rplC against the tested RGM. A1la77Pro mutation was detected in rplD which is located in variable site and is far away from LZD-binding site, as shown by the sequence alignment. Except A2271G mutation in 23SrRNA in M. massiliense that was closer to binding site of LZD, other mutations are far from the LZD-binding site. Our results combining MIC test, gene mutation and structure based analysis showed there was no obvious correlation between riboproteins mutations(rplC and rplD)and LZD resistance against tested RGM species. Mutations located in the LZD binding site may cause LZD resistant. Hence, rplC, rplD and 23srRNA homologues might not be the only target for LZD to explore its bacteriostatic activity.
In conclusion, this study demonstrated that oxazolidinones have good in vitro activities against the overwhelming majority of RGM species. The efficacies of the four oxazolidinones were variable against different species. TZD showed strongest antimicrobial activity against M. abscessus and M. massiliense, while DZD owned the strongest activity against M. fortuitum. The data provided important insights into the possible clinical applications of oxazolidinones to treating RGM infections(39).
{kind=link}
{kind=link}