Clostridioides difficile infection is a particularly interesting ecosystem to explore for this task, since disruption of the gut microbiome ecosystem by antibiotics is a major risk factor for the condition, and ecosystem restoration by FMT is a well-established treatment for the condition. In a previous longitudinal multi-omics study9, faecal samples from human donors were implanted in artificial gut models (Fig. 1) to simulate the gut environment in vitro to study the effect of FMT on inhibiting the growth of C. difficile. Samples were collected from the models on alternate days, and bacterial abundance (16S rRNA gene sequencing), bile acid and metabolite profiles were generated from these samples, which were called target variables in the analysis.
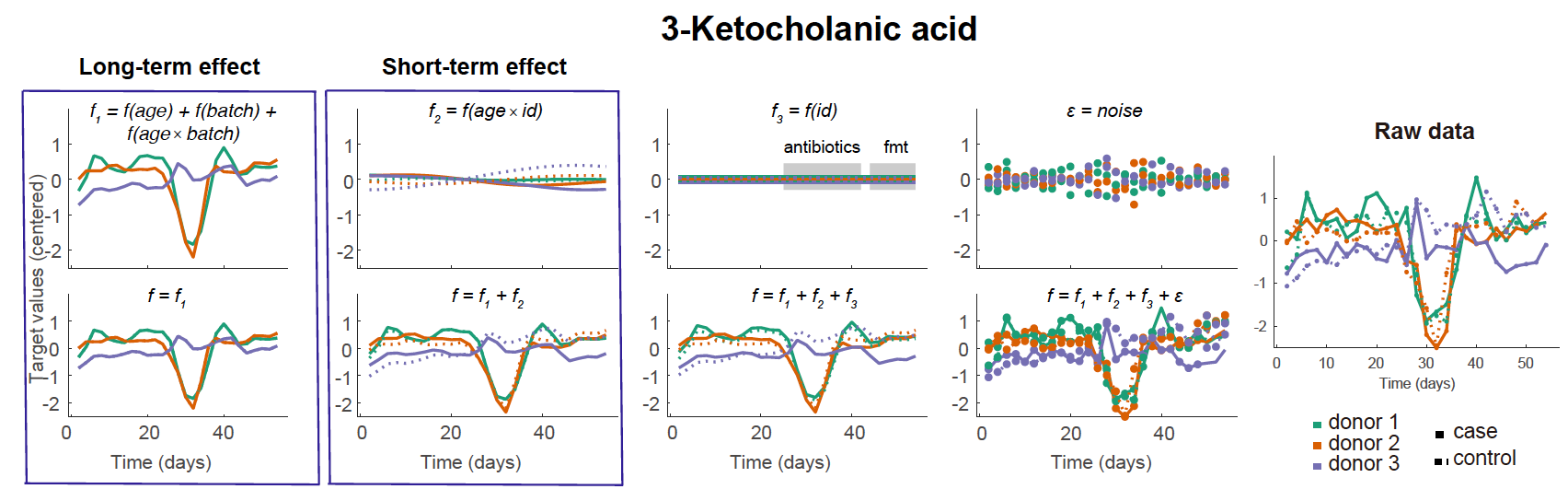
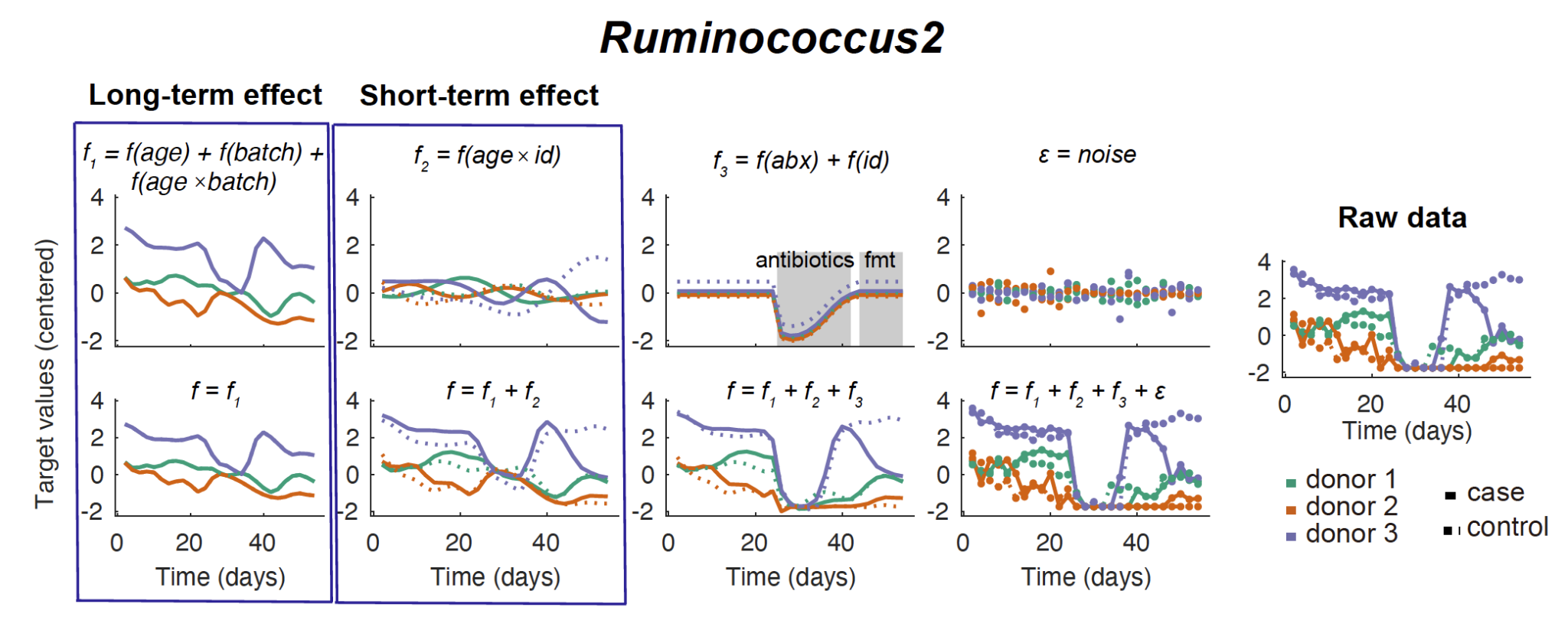
We first used LonGP7 to deconvolute each target variable into various covariate associated components, such as the global trend shared by all models, the batch specific trend shared by models that use faecal samples from the same donor, the FMT trend shared by cases and short-term trend that is model-specific. This analysis has successfully uncovered that valerate concentration first decreases after the introduction of antibiotics and only increases after FMT (Fig. 2a), which confirms the serendipitious main finding of the original publication and helps to validate our approach.
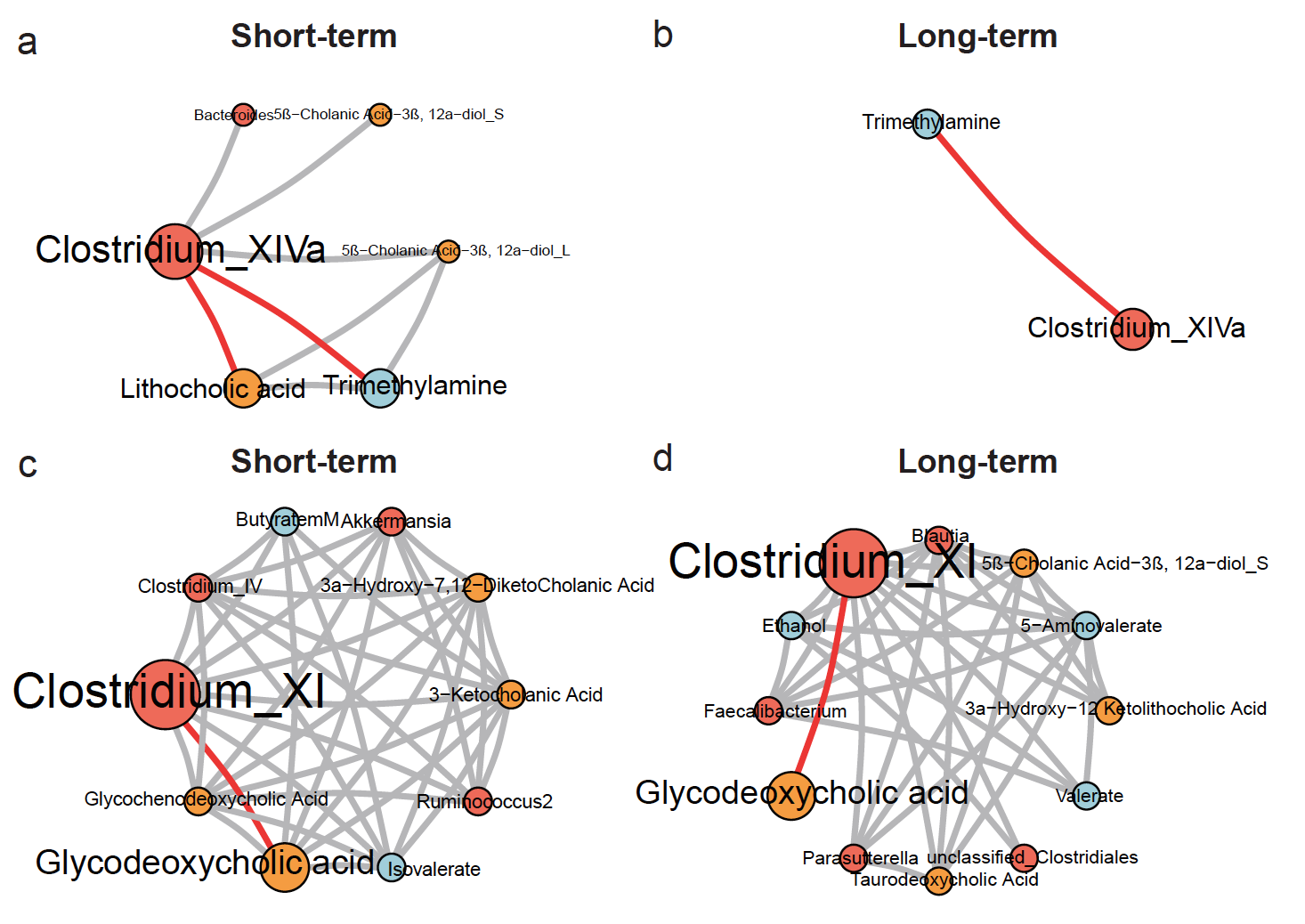
Based on the LonGP regression results, we extracted the long-term and short-term trends to construct separate bacteria-metabolite interaction networks using WGCNA11. The long-term interaction network (Fig. 2b, Supplementary Data 3) led us to focus on Blautia, which was not involved in any interaction in the raw data interaction network (Supplementary Data 3). These negative interactions (Blautia with ethanol, 5-aminovalerate and succinate) were presumed to be weak and hard to detect using direct associations. Batch culture experiments incubating Blautia obeum in brain-heart infusion (BHI) medium validated the interaction with succinate (Fig. 2c), but not interactions with ethanol and 5-aminovalerate (no change in all replicates). This interaction between Blautia and succinate would not have been expected without the network analysis, and to our knowledge has not been previously-described.
The short-term interaction network (Fig. 2d, Supplementary Data 3) identified new transient interactions compared with the raw data network (Fig. 2e), e.g. Ruminococcus2 positively correlate with 3-ketocholanic acid (KCA), whose LonGP deconvolutions were provided in Supplementary Fig. 1–2. In batch culture experiments (Fig. 2f) incubating two representative human microbiome members of Ruminococcus2 (R. gnavus and R. luti) with lithocholic acid (LCA), we found that KCA was generated by both R. gnavus and R. luti. In particular, KCA was detected in the batch culture of R. gnavus in media with LCA at baseline, which is surprisingly fast. This was not contamination since the LCA control (identical medium without R. gnavus) contained no KCA, which suggested KCA must be generated during the set up of the batch culture at 0 hour (took ~ 10 minutes). Reduced KCA was detected in the R. gnavus + LCA batch culture at 24 hours compared to 10 minutes, suggesting further ongoing bacterial metabolism. No KCA was detected in the baseline R. luti/ LCA batch culture but was detectable by 24 hrs; this suggests variability in the dynamics between Ruminococcus2 members at which LCA is metabolised to KCA.
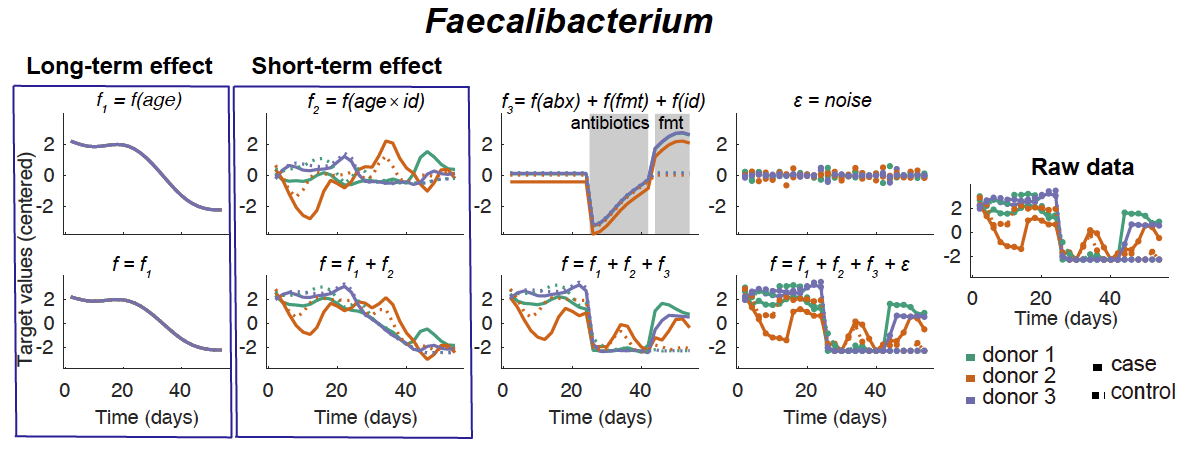
Other findings from the networks supported established knowledge about bacteria-metabolite interactions; for example, a positive correlation was observed between the genus Clostridium XIVa and lithocholic acid in the short-term network (Supplementary Fig. 3a), with this being one of the only genera containing bacteria able to bioconvert primary to secondary bile acids17. The placement of trimethylamine (TMA) in short- and long-term networks (Supplementary Fig. 3a,b) is supported by the prior recognition that TMA is one of the identified metabolites most strongly restored by FMT for C. difficile infection1 (CDI). Similarly, the positive interaction between glycodeoxycholic acid (GDCA) and Clostridium XI (the genus containing C. difficile) seen in short and long-term networks (Supplementary Fig. 3c,d) supports the prior recognition of GDCA in promoting germination of spore forming bacteria including C. difficile18. A further notable network feature is the loss of Faecalibacterium after antibiotics and restoration after FMT (Supplementary Fig. 4), especially of note given the previous observation of the absence of F. prausnitzii in inflammatory intestinal conditions including CDI and inflammatory bowel disease19, and the anti-inflammatory properties associated with this bacterium.
{kind=link}
{kind=link}
{kind=link}
{kind=link}