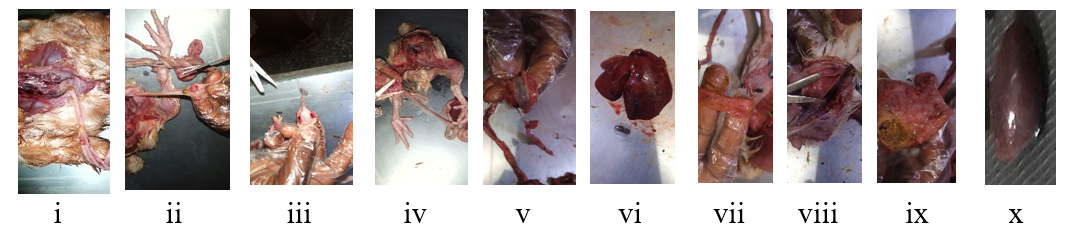
Characteristic pathological lesions on organs seen during post mortem examination
Molecular detection of NDV
The overall M-gene detection rate by RT-PCR was 54% (27/50). A 121-bp fragment was amplified and products of electrophoresis visualized by Ultraviolet (UV) trans-illumination (Fig 1).
F-gene amplification was successfully carried out in seven samples and products of electrophoresis visualized by UV trans-illumination (Fig 2,3). 1662bp nucleotide sequences obtained were deposited in the GenBank and accession numbers assigned (Tab 2).
Tab 2: Sample collected, Sequence ID and corresponding Accession Number
|
Sample collection ID
|
Sequence ID
|
Isolate specimen voucher
|
Isolated From
|
Accession Number
|
|
PCR/014/200320
|
Seq 14
|
F gene KN 14
|
Broiler
|
OK491971
|
|
PCR/036/250320
|
Seq 36
|
F gene KN 36
|
Broiler
|
OK491972
|
|
PCR/048/300320
|
Seq 48
|
F gene KN 48
|
Broiler
|
OK491973
|
|
PCR/055/310320
|
Seq 55
|
F gene KN 55
|
Broiler
|
OK491974
|
|
PCR/056/310320
|
Seq 56
|
F gene KN 56
|
Broiler
|
OK491975
|
|
PCR/071/020420
|
Seq 71
|
F gene KN 71
|
Layer
|
OK491976
|
|
PCR/075/030420
|
Seq 75
|
F gene KN 75
|
Layer
|
OK491977
|
Phylogenetic tree and evolutionary distance analysis
Phylogenetic tree was constructed (Fig 4) using MEGA (11.0). The genetic relatedness of the isolates, a Lasota reference strain KU665482 and other NDV genotypes of known F gene sequences obtained from GenBank database was inferred by phylogenetic analysis. All isolates clustered around the newly classified genotype XIV (sub genotype XIVb) in class II which is widely reported in Nigeria.
Nucleotide blast analysis shows a 99% nucleotide identity to virulent NDV MT543153 isolated in 2019 from backyard poultry in Niger (a country to the North border of Nigeria). KY171993 isolated from Nigeria in 2009 shows a homology 97.65% (as at 16th February, 2022; data not shown). The nucleotide sequence homologies between these isolates and the common vaccine strains used in the country is shown (Tab 3).
Tab 3: Estimates of evolutionary divergence and percentage homology between study isolates and commonly used vaccines in Nigeria
|
|
KU665482 NDV LaSota straina
|
KT445901 NDV Komorov straina
|
|
Isolatea
|
Genetic distanceb
|
% Homologyc
|
Genetic distanceb
|
% Homologyc
|
|
OK491971
|
0.23
|
81.44
|
0.22
|
81.86
|
|
OK491972
|
0.22
|
81.56
|
0.22
|
81.98
|
|
OK491973
|
0.23
|
81.50
|
0.22
|
82.04
|
|
OK491974
|
0.23
|
81.44
|
0.22
|
81.86
|
|
OK491975
|
0.29
|
81.44
|
0.22
|
81.86
|
|
OK491976
|
0.23
|
81.26
|
0.22
|
81.76
|
|
OK491977
|
0.23
|
81.20
|
0.22
|
81.65
|
acalculated from the complete F gene sequences
bThe number of base substitutions per site from between sequences are shown. Analyses were conducted using the Maximum Composite Likelihood model [17]. This analysis involved 9 nucleotide sequences. Codon positions included were 1st+2nd+3rd+Noncoding. All positions with less than 95% site coverage were eliminated, i.e., fewer than 5% alignment gaps, missing data, and ambiguous bases were allowed at any position (partial deletion option). There were a total of 1662 positions in the final dataset. Evolutionary analyses were conducted in MEGA11 [18]
c Align sequence nucleotide blast showing homology of a 99% query cover on NCBI blastn suite
Molecular Characterization and Mutational Analysis of the Functional and Antigenic domains
The complete translated 553 fusion protein amino acid sequences obtained from the study isolates was used to compare their functional and antigenic domains relative to LaSota vaccine strain. Notable substitutions around these regions were observed. Interestingly, among the research isolates, these substitutions were sometimes not observed similarly. Numbering system of aa was used to name the detected aa substitutions with respect to observed genetic variations.
Along the hypervariable region (residue 1-31), 14 substitutions were observed (Fig 5). In comparison to the La Sota reference strain KU665482, all isolates displayed a S31P in the signal peptide region as a result of TCC to CCC substitution at the 91-93. This mutation was seen only in KJ958914 and KJ958913 (obtained from GenBank) among the 69 isolates included during the alignment and phylogenetic study (supplementary document). Isolate OK491976 exhibited unique aa substitutions AGA to GAA, GCA to GAA and CTG to CAG at codons 10-12, 31-33 and 43-45 respectively leading to Arginine to Glutamate at position 4, Alanine to Glutamate at position 11 and Proline to Glutamine at position 15. In contrary, all other study isolates exhibited AGA to AAA and GCA to GTA substitution at codons 10-12 and 31-33 leading to Arginine to Lysine at position 4 and Alanine to Valine substitution at position 11 respectively while the proline at position 15 is conserved. P28 is conserved in OK491972 but not in the other six isolates with CCG to CTG substitution at codons 82-84 leading to Proline to Leucine substitution. A29 was conserved in OK491971 and OK491975 but substituted in the other research isolates due to GCA to ACA substitution at codons 85-87 leading to Alanine to Threonine substitution.
Eight transmembrane domains have been reported [19] at residues 14-27, 15-25, 118-131, 120-128, 266-269, 429-432, 499-525 and 501-523. Compared to the LaSota strain, this region is highly non-conserved in all isolates except for 266-269 with no amino acid substitution (Fig 5,6).
All seven isolates share the characteristic virulent motif 112R-R-R-K-R/F117 at the F0 cleavage site indicating that they are velogenic NDV strains (Fig 5). GGC to AGA substitutions at codons 334-336 causing a Glycine to Arginine substitution at position 112; there was AGA to CGA substitution at codon 337-339 in all isolates though Arginine residue is present in both LaSota and research isolates at position 113; CAG to CGG substitution in codon position 340-342 causing a Glutamine to Arginine substitution at position 114; Glutamine to Lysine substitution at position 115 occurred in all isolates, however, isolates OK491973 and OK491976 exhibit GGC to AAA substitution at codons 343-345 while the others show GGC to AAG substitution. At position 116, both LaSota and research isolates exhibit Arginine residue however, aa CGC and CGT is present at codon 346-348 in Lasota and research isolates respectively. CTT to TTT substitution at codon 349-351 occur leading to Leucine to Phenylalanine substitution at position 117 in all the isolates.
Along the fusion peptide region (117-142), five aa substitutions is seen. While L117F and I118V is expected in the virulent furin-like molecule, the I121V, G124S and I135V substitutions are not. This substitutions is due to GGT to AGT nt substitution at codon 361-363; GGT to AGT substitution at codon 370-372 and ATA to GTA substitution at codon 403-405 respectively.
In addition, the F protein has six highly conserved potential N-linked glycosylation sites Ng1-Ng6 [20] with sequence Asn(Asparagine)-X-Ser(Serine)/Thr(Threonine) (N-X-S/T) where X is any aa except proline and aspartate [19,21]. Amino acids at these sites were used and conserved in all NDV isolates of this research at residue 85NRT, 191NNT, 366NTS, 447NIS, 471NNS and 541NNT. Hence, no loss of glycosylation site though there was one substitution compared to the LaSota strain at residue 191NKT. In addition, same sense point mutations occurred at some points as; Asparagine at residue 85 and Arginine at residue 86 show nt mutation but no aa mutation from AAC to AAT (Asparagine) and AGG to AGA (Threonine) in isolates compared to LaSota strain in codon 253-255 and 256-258 respectively. Similarly, at codon 1096-1098, nt substitution from AAT to AAC is observed only in one isolate OK491977 and TCG to TCA substitutions in all isolates at codon 1102-1104, nevertheless, both substitutions did not alter the aa residue Asparagine and Serine at positions 366 and 368 respectively. Asparagine residue at 472 exhibited nt substitution AAC to AAT at codon 1414-1416 while the Serine residue at 473 showed nt substitution only in one isolate OK491977 from TCG to TCA. There was nt substitution from AAT to AAC in the codon 571-573 of the Asparagine residue at position 191. AAA to AAT nt substitution at codon 574-576 cause a Lysine to Asparagine substitution at residue 192. There is nt substitution from ACA to ACG at codon 577-579 of the Threonine residue at 193. All positions 447NIS were all conserved with no nt substitution compared to LaSota. Lastly, the Threonine residue at 543 show nt substitution from ACT to ACC.
Cysteine residues are important in the connection between F1 and F2 subunits. The conserved cysteine in the C-terminal has been reported to maintain the F protein structure. Cysteine residues conserved at positions 25, 27, 76, 199, 338, 347, 362, 370, 394, 399, 401, 424, 514 and 523 of the F protein in most NDV isolates [21]. Amino acids are used and conserved in all the isolates except for a unique point cysteine (C) to Serine substitution at residue 394 in OK491977. No nt substitution was observed in the Cysteine residues at position 25 (codon 73-75), 76 (codon 226-228), 199 (codon 595-597), 347 (codon 1039-1041), 370 (codon 1108-1110), 399 (codon 1195-1197), 401 (codon 1201-1203), 424 (codon 1270-1272), 514 (codon 1540-1542) and 523 (codon 1567-1569). Nucleotide substitutions was observed at codon 79-81from TCT to TGC of the Cysteine residue at position 27 only in isolate OK491974, nt substitution of cysteine residue 338 from TGT to TGC at codon 1012-1014, TGC to TGT substitution at codon 1084-1086 at Cysteine residue 362, nt substitution at Cysteine residue 394 in OK491977 from TGC to AGT at codon 1180-1182 resulted in a Cysteine to Serine substitution while TGC to TGT (Cysteine) was observed in all the other isolates. Therefore one Cysteine residue site is lost in isolate OK491977.
The major epitopes involved in virus neutralization are conserved in all residues except for one amino acid substitution Lys AAG to Arg AGA (K78R) of the A2 neutralizing epitope identified in all isolates (Fig 5). However, nt substitution occurred even in the conserved epitopes as; Aspartate residue on position 72 show nt substitution from GAT to GAC at codon 214-216 in all isolates; Glutamate residue on position 74 show nt substitution from GAG to GAA at codon 220-222 in all isolates; Alanine residue on position 75 show nt substitution from GCA to GCG at codon 223-225 in all isolates; there was nt substitution from AAA to AGA at codon 232-234 in all isolated that lead Lysine to Arginine amino acid substitution in all isolates and the loss of the A2 neutralizing epitope; at codon 235-237, there is no nt substitution in the Alanine residue. Only 4 nt substitutions is observed along the antigenic epitope 151-171; codon 457-459 nt substitution from CGA to CGG of the 153-Arginine residue; codon 463-465 substitution from AAA to AAG of the 155-Lysine residue; codon 487-489 substitution from GAG to GAA of the 163-Glutamate residue; codon 505-507 is conserved in isolate OK491975 but nt substitution is seen in the others from ACT to ACC of the 169-Threonine residue.
The three Heptad Repeat regions HRa (143-185), HRb (268-299) and HRc (471-500) in the isolates displayed 1, 5 and 6 aa substitutions respectively compared to the LaSota reference strain. K145N at HRa; N272Y, S278P, I285K, T288N, N297K at HRb and N476T, N479D, E482A, R486N, K494R and T498S. Notably, substitutions I285K was seen only in OK491973 and OK491975 only. N297K was seen only in isolate OK491973 only. N476T was observed only in isolate OK491976. Interestingly, T498S was observed in all research isolates but not in any other isolate included in the phylogenetic tree analysis even those isolated previously from Nigeria or Africa (Fig 6, supplementary material).
There is 14 nt substitutions along the HRa region; GCC to GCT at codon 430-432 of the 144-Alanine residue; AAA to AAT (OK491971 only) and AAA to AAC in codon 433-435 lead to Lysine to Asparagine substitution at residue 145; CAA to CAG nt substitution in all isolates at codon 436-438 of 146-Glutamine residue; GCT to GCC substitution at codon 442-444 of 148-Alanine residue; GCA to GCT nt substitution at codon 445-447 of Alanine-149 in isolates OK491945 and OK491947 only; AAC to AAT substitution at codon 448-450 of 150-Asparagine in OK491971, OK491972, OK491974, OK491975 and OK491977 only), CGA to CGG nt substitution is seen in all isolates at codon 457-459 of 153-Arginine; CTT to CTC at codon 460-462 of the 154-Leucine residue; AAA to AAG nt substitution in codon 463-465 of the 155-Lysine residue; GAG to GAA nt substitution in 487-489 of 163-Glutamate residue; ACT to ACC nt substitution of codon 505-507 of 169-Threonine residue; TCG to TCA nt substitution in codon 517-519 of 173-Serine residue in isolates OK491974 and OK491975 only; GCG to GCA in codon 532-534 of 178-Alanine residue and GTT to GTC in codon 535-537 of 179-Valine residue is seen in all isolates.
Eighteen nt substitutions occur along the HRb region; TTA-TTG in codon 802-804 of the 268-Leucine residue; AAC-TAC in codon 814-816 that cause aa substitution Asparagine to Tyrosine at residue 272; CCT-CCC nt substitution in codon 817-819 of the 273-Proline residue; ATT-ATA nt substitution in codon 820-822 of the 274-Isoleucine residue; CTA-CTG nt substitution in codon 823-825 of 275-Leucine residue; TAC to TAT nt substitution in the codon 826-828 in the 276-Tyrosine residue; TCA to CCA nt substitution in codon 832-834 cause aa substitution form Serine to Proline at position 278; CAG to CAA nt substitution in the codon 835-837 of 279-Glutamine is seen only in isolates OK491973 and OK491976; CAA to CAG substitution in codon 841-843 of the 281-Glutamine residue is seen in OK491971, OK491972 and OK491976 only; GGT to GGC nt substitution in codon 850-852 in the 284-Glycine residue; ATA to AAA nt substitution in codon 853-855 is seen in OK491973 and OK491975 only resulting to aa substitution from Isoleucine to Lysine at position 285; CAG to CAA nt substitution in codon 856-858 in the 286-Glutamine residue in all isolates except OK491973; ACT to AAT nt substitution in codon 862-864 cause aa substitution from Threonine to Asparagine at position 288; CTA to TTA nt substitution is seen in codon 865-867 in the 289-Leucine residue; CCT to CCC in the codon 868-870 of the 290-Proline residue; TCA to TCG nt substitution in the codon 871-873 of the 291-Serine residue is seen in all the isolates except OK491974; GTC to GTT in the codon 874-876 of the 292-Valine residue; AAT to AAC nt substitution is seen in the codon 889-891 in 297-Asparagine in OK491974, OK491976 and OK491977 only, whereas, AAT-AAA nt substitution in the same position is seen in OK491973 only causing Asparagine to Lysine aa substitution in position 297.
Along the HRc region, there are 17nt substitution. AAC-AAT nt substitution in the 1414-1416 codon of the 472-Asparagine residue; TGC to TCA nt substitution in the 1417-1419 codon in the 473-Serine residue in isolate OK491977 only; ATC to ATA and AGT to AGC nt substitution occur in the 474-Isoleucine and 475-Serine residues respectively; AAT to ACT nt substitution in the 1426-1428 is seen in isolate OK491976 only causing Asparagine to Threonine aa substitution at position 476; TTG to CTG nt substitution occur in the 1432-1434 478-Leucine residue; AAT to GAC in codon 1435-1437 cause aa Asparagine to Aspartic acid substitution at position 479; TTA to TTG nt substitution in the 1441-1443 in the 481-leucine residue; in codon 1444-1446, GAG to GCA cause Glutamic acid to Alanine aa substitution at position 482; AGC to AGT nt substitution in the 1450-1452 of 483-Glutamic acid residue; AGA to AAC nt substitution in codon 1456-1458 cause aa substitution Arginine to Asparagine in position 486; AAA to AAG nt substitution in the 1459-1461 codon of the 487-Lysine residue; AAA to AGA nt substitution in the 1480-1482 codon cause aa Lysine to Arginine substitution in position 494; CTG to CTA, ACT to AACC, AGC to AGT nt substitutions in the codon 1483-1485, 1486-1488 and 1489-1491 of the 495-Leucine, 496-Threonine and 497-Serine; ACA to TCA substitution in the 1492-1494 codon cause aa Threonine to Serine at position 498.
There are thirteen effective B-cell epitopes for LaSota vaccine as surface-exposed amino acids which were speculated to cause antigenic difference between the vaccine and wild strains [19]. Amino acid sequence analysis of the isolates revealed multiple substitutions within these regions (Fig 4).
{kind=link}