Pacific bluefin tuna (PBT) Thunnus orientails (Temminck & Schlegel) is regarded as a superior foodstuff and is used for sushi and sashimi, mainly in Japan. Consequently, this is a fish species with extremely high commercial value. Therefore, PBT have been caught internationally by a variety of fishers, and consequently, their population levels have remained low for decades (Nakatsuka et al. 2017). Successful PBT aquaculture could reduce fishing pressure and provide a stable supply of fish. However, in recent years, even in Japan, more than half of the young fish used for aquaculture depended on natural resources (Sawada et al. 2005).
Several institutions continue to research seedling production in T. orientalis, including Kinki University where PBT was successfully completely farm-raised for the first time in the world, Unfortunately, translating this into mass production is difficult. The high mortality in the larval and juvenile stages (Harada et al. 1971; Miyashita 2002; Sawada et al 2005; Ishibashi 2012) and consequent required numbers of fish prevent the process from being profitable. The number of fish larvae was depleted as a result of floating and sinking deaths in pre-flexion larvae, and cannibalism in post-flexion larvae to juveniles (Sawada et al. 2005; Ishibashi et al. 2008; Sabate et al. 2010; Tanaka et al. 2018). In the juvenile stage, cannibalism, and mass death occurs due to fractures and skin lacerations caused by contact with rearing tanks and offshore net cages (Ishibashi 2012; Okada et al. 2020a, b). However, there are effective approaches to instances of mass death in PBT such as the addition of feed oil to prevent floating deaths (Kurata et al. 2012), and 24-hour lighting in the rearing environment to prevent sinking deaths (Kurata et al. 2017). Furthermore, to prevent deaths from cannibalism, we ensured that the fish were of uniform size and adequately fed (Ishibashi et al. 2013a). Deaths caused by fractures and skin lacerations can be prevented by using particular colours and patterns in the rearing tanks (Ishibashi et al. 2013b; Okada et al. 2015), and providing light at night in the sea net cages (Ishibashi et al. 2009; Honryo et al. 2020). Although several research and private institutions have introduced these technologies into PBT seedling production, their production efficiency is still low compared with that of other marine fish.
Seedling production is carried out on land and the size of the facility cannot easily be increased, owing to restrictions such as the size of the land and running costs. Furthermore, PBT is a multiple-spawning fish, with a spawning season that is limited to approximately 60 days along the coast of Wakayama Prefecture (Miyashita 2002). However, it takes approximately 30 to 40 days after the eggs are fertilized in the production facility tanks for them to be raised to juveniles ready for transfer to the marine cages (Miyashita 2002). Consequently, it is not possible to have many production rounds. Therefore, seedlings have to be produced at a higher stocking density to increase the production efficiency. However, PBT larvae have been empirically bred at a much lower density than that of other marine fish. Chasing behaviour is observed in PBT larvae after the post-flexion stage (Sabate et al. 2010), and, hatched larvae of other fish species are fed as live baits (Sawada et al. 2005). Therefore, mass death due to cannibalism is a concern (Ishibashi 2012; Tanaka et al. 2018).
Cannibalism is a serious problem in seedling production (Baras and Jobling 2002) and is an important factor for survival. This phenomenon is accompanied by aggressive behaviour as a precursor and is affected by the rearing environment. For example: in Koi carp (Cyprinus carpio) and Asian seabass (Lates calcarifer) cannibalism is promoted by high stocking density (Damme et al. 1989; Khan et al. 2021) while differences in fish size promotes it in the orange-spotted grouper (Epinephelus coioides), (Hseu 2002). In yellowtails (Seriola quinquevadiata) and greater amberjack (Seriola dumerili), differences in fish size and starvation increased the frequency of aggressive behaviour, but a higher stocking density decreased it (Sakakura and Tsukamoto 1998; Miki et al. 2011). Stocking densities that are too high can stress fish and adversely affect their survival and growth (Barton and Iwama 1991).
Many fish species, such as the gilthead sea-bream (Sparus aurata), Atlantic cod (Gadus morhua), and Nile tilapia (Oreochromis nilioticus), slow down growth as stocking density increases (Canario et al. 1998; Lambert and Dutil 2001; El-Sayed 2002). Conversely, increasing stocking density promotes growth in Arctic char (Salveelinus alpinus) (Wallace et al. 1988), Atlantic Salmon (Salmo salar) and European perch (Perca fluviatilis) had no effect on growth (Kjartansson et al. 1988; Baras 2003). It is necessary to take note of the developmental stage and the degree of stocking density of the test fish used in each experiment to make comparisons, but whether the increased stocking density affects survival and growth depends on the fish species.
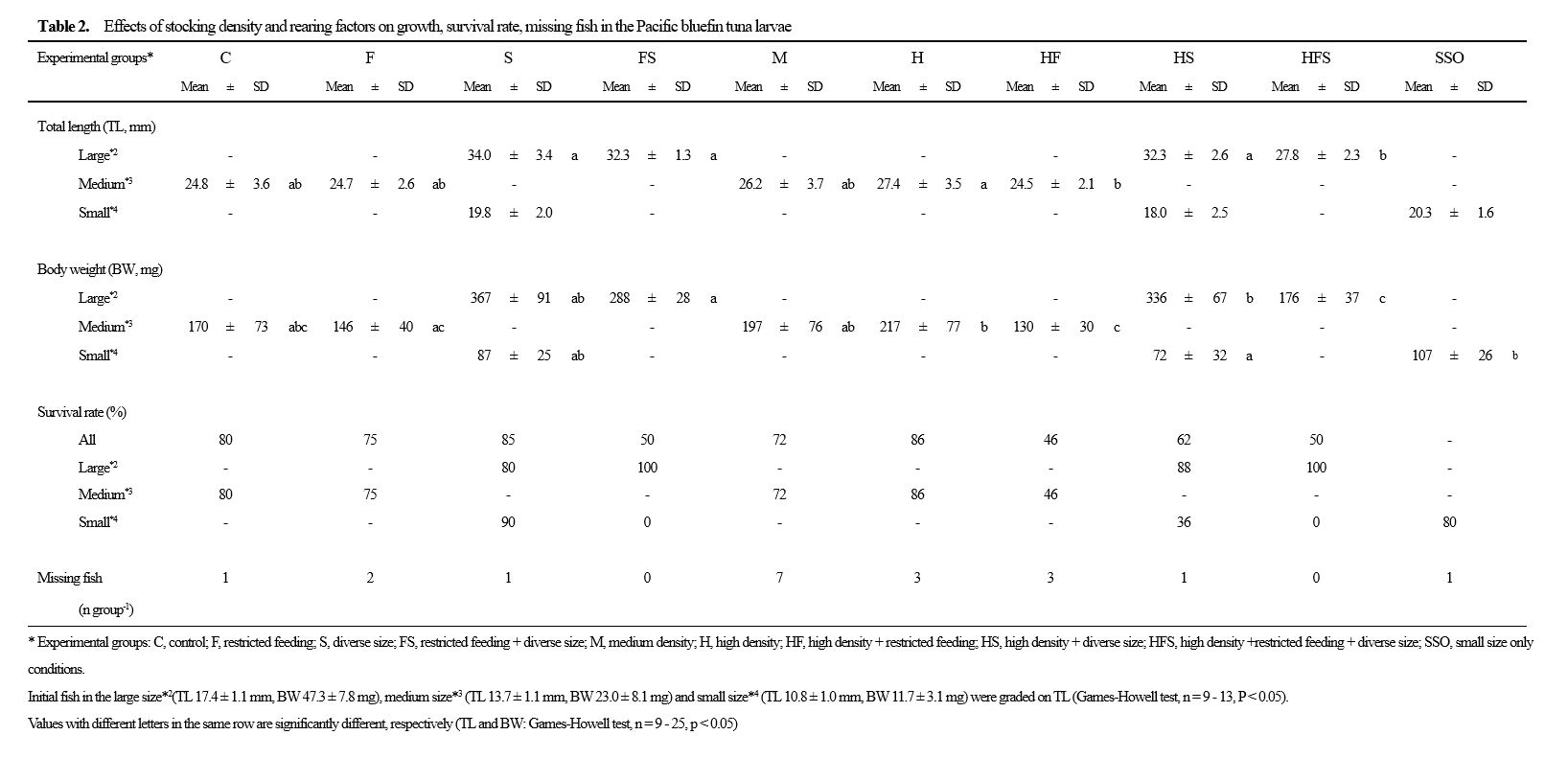
In PBT larvae, aggressive behaviour have been shown to increase by feeding is restricted, and survival rates are reported to deteriorate further when size differences occur (Ishibashi et al. 2013a). Therefore, it is possible that PBT larvae can be produced at a higher density by feeding them sufficiently and selecting and rearing them such that the size difference is small. However, the effects of stocking density on aggressive PBT behaviour and cannibalism is not well understood. In this study, to ensure sufficient production and reduce the unit price per seedling, we paid particular attention to the stocking density of larvae in the post-flexion stage, where mass deaths due to cannibalism is a concern. PBT larvae were reared at various densities, and the effects of stocking density on aggressive behaviour, cannibalism, survival and growth were investigated. Moreover, factors such as feeding restrictions and fish size differences in addition to stocking density were considered to have a significant effect on the aggressive behaviour, cannibalism, survival and growth of PBT larvae (Ishibashi et al. 2013a).
{kind=link}