In this study, we report that mice lacking progesterone receptor signaling in the osteoprogenitor cells were more susceptible to collagen-induced arthritis, especially male mice. The male PRΔPrx1 -CIA mice had a significantly higher incidence of arthritis, joint inflammation, bone erosion, and cartilage damage compared to the male PRΔPrx1 mice or WT-CIA mice. Our findings indicate that under "normal" conditions, the presence of PR in osteoprogenitor cells might be protective for rheumatoid or inflammatory arthritis incidence and might also partially account for the sex differences that are observed in RA patients [53–55].
Some susceptibility genes for RA have been previously identified. The human leukocyte antigen (HLA) is a genetic site controlling immune responses in RA [56, 57]. Several genes outside the HLA region, such as Stat4, the TRAF1-C5 locus, and PTPN22, have been reported to be associated with activation and progression of inflammation in RA [58–62]. There is hardly any information explaining the sex disparities in genetic susceptibility to RA except a polymorphism in the Cyb5a gene, which is related to androgen synthesis, was found to be associated with risk for RA in women but not in men [63]. Recent studies have also suggested a role for epigenetic modifications in the activation and aggressiveness of synovial fibroblasts in RA [64–67] and the X-encoded genes, Timp1 and IL-9R also may play a role [68]. Some of these epigenetic modifications have been linked to X-linked miRNA, and the presence of the second X chromosome in females may affect miRNA expression levels that could account for the development of sex-related autoimmunity [69, 70]. Most of these studies on sex-specific factors affecting RA have focused on the potential effects of sex hormones due to the observation that RA improves during pregnancy and that male RA patients generally have a less severe course of illness and better response to therapy [5, 9, 10]. Estrogen has been reported to have both pro-inflammatory and anti-inflammatory effects on the immune system while both progesterone and androgen are anti-inflammatory [11–19]. The effects of hormones are mainly regulated through their hormonal receptors. The presence and proportion of estrogen and androgen receptors in different tissues and cells, including fibroblasts, chondrocytes, and bone cells, might define their roles in the sexually-dimorphic pathogenesis of RA [20–27]. We and others have found PR expressed in growth plate chondrocytes, osteoclasts, and osteoblasts, and PR plays a critical role in peak bone mass determination [37, 71, 72]. We also have reported that the loss of PR signaling in osteoprogenitor cells regulates key signaling pathways for immune response, especially in males [34]. We identified PR-targeted genes that regulated sex differences, including an "X-inactive specific transcript," Xist, Mtus2, Aldhla7/1, Tusc5, Cd300c, and Pde3a [34]. The upregulation of Xist is associated with chronic inflammation and pain in females with complex regional pain syndrome [73] and contributes to RA progression [74]. Cd300c and Pde3a were over-presented in RA patients [75, 76] and were associated with inhibition of T cell immunity [77] or response to TNF inhibitors in RA patients [78]. Taken all together, PR appears to regulate susceptibility to RA in mice, targeting several genes that regulate sex differences and which might correlate with clinical observations of sex-differences in RA.
The presence of marginal bone erosions, detected by imaging, predicts a more severe disease course with more disability and increased morbidity. The significance of erosions in RA has been the focus of the development and approval of several agents for modifying the course of RA and has been validated in clinical trials as being able to reduce structural joint damage, including bone erosion and cartilage degradation [79, 80]. The bone erosions in RA show a predilection for specific anatomic sites such as the radial aspects of finger joints, while the ulnar aspects are relatively spared [81]. These focal erosions typically emerge at the site at which the synovium comes into direct contact with the bone which is known as bare areas. Anatomical factors that predispose these skeletal sites for erosion include the presence of mineralized cartilage, the insertion of ligaments at the bone surface, and inflamed tendon sheaths that enable the spread of inflammation from the tendon to the articular synovium. Articular erosion at these “bare areas” represent localized bone loss from osteolysis, which resulted from an imbalance in which bone resorption by osteoclasts is predominant over bone formation by osteoblasts. Once established, these bone erosions rarely repair despite the use of potent biologic therapeutic strategies including biologics such as TNF, IL-1 or IL-6 receptor blockade [82–85]. Aberrant repair of erosions appeared as sclerosis with new bone apposition at the base of the erosion and might involve the juxta-articular bone marrow. Adipose tissue might populate the erosive area. Bone erosion seemed to correlate with on-going inflammation. Our study provides additional information to better understand the potential PR regulation of the inflammation-bone resorption coupling mechanism in the process of joint and bone damage, and how potentiation of this coupling from lack of PR signaling contributes to bone and joint loss in RA in a sex-dependent manner.
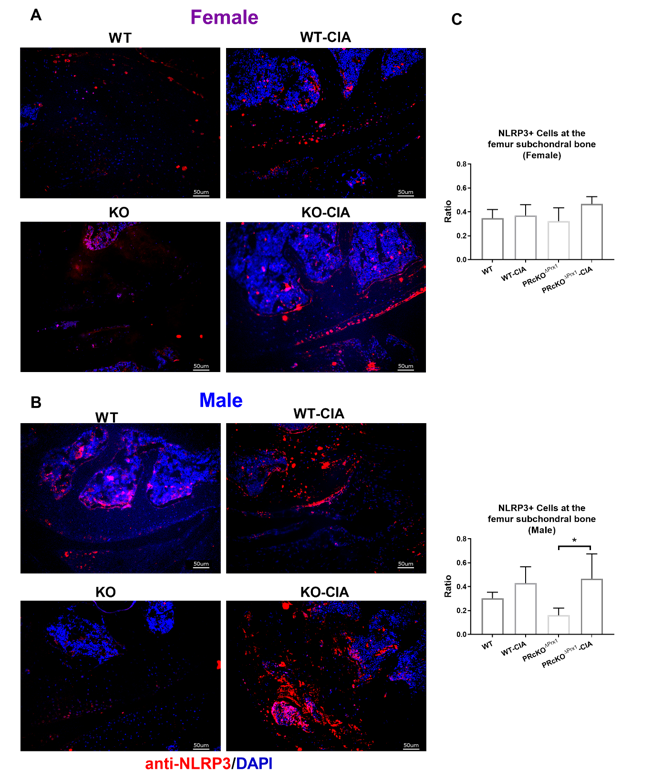
Recent studies have reported that inflammasomes contribute to the pathogenesis of several age-related metabolic diseases, including atherosclerosis [86–89] and Alzheimer’s disease [90–95]. Inflammasome activation resulting in IL-1β production is an essential part of the innate immune response. NLRP3 binds with the adaptor apoptosis-associated speck-like protein (ASC) and triggers the cleavage/activation of pro-caspase-1, which catalyzes the activation of IL-1β, contributing to the progression of inflammation [96]. Genetic variants within the NLRP3-inflammasome are associated with susceptibility to RA [97–101], and enhanced activity of NLRP3 inflammasome has been found in peripheral blood cells of patients with active RA [102]. Inhibition of NLRP3 inflammasome activity alleviated arthritis in CIA mice [103]. Given that NLRP3 was one of the PR-targeted gene [34] and that the lack of PR potentiated NLRP3 inflammasome expression in the synovial tissue of the CIA mice, suggesting that although PR might not alter the underlying cause of disease, it might reduce the production of factors such as inflammasomes which are involved in inflammation, cartilage and bone destructions in RA.
In conclusion, we found that the lack of PR in osteoprogenitors increased susceptibility to IA, especially in male mice. Our findings indicate that the presence of PR in osteoprogenitor cells counteracts the development of collagen-induced arthritis and might also help to explain sex differences observed in rheumatoid arthritis.
{kind=link}