M. oryzae-infected MoSDT1-transgenic rice showed accumulation of multiple defensive metabolites and activation of multiple metabolic pathways
Previous studies revealed that when pathogens infect plants, the different metabolites induced by the infection play an essential role in strengthening the plant resistance and alleviate the disease symptoms. Our prior study confirmed that the heterologous expression of MoSDT1 in rice increases tiller numbers without affecting its nuclear morphology. Most importantly, it also resulted in a significant increase in resistance to M. oryzae in rice, mainly presented with early induction of massive callose deposition, ROS accumulation, cell death, and regulation of defense-related gene expression in infected MoSDT1-transgenic rice. Moreover, specific primary metabolites that participate in defense response accumulated in MoSDT1-transgenic rice [18]. To further investigate the biochemical mechanism of the enhanced resistance in M. oryzae-infected MoSDT1-transgenic rice, the metabolomics was analyzed to better explore the core metabolites and critical metabolic pathways of M. oryzae-infected MoSDT1-transgenic rice.
Organic oxygen compounds, organic acids, enzymes, and glycoside are the first category of metabolites produced by infected plants [2]. The accumulation of organic oxygen compounds such as sucrose, raffinose, galactinol, D-mannose, and maltotriose was identified at 0 h, 72 h, and 120 h in this study. D-mannose is mainly involved in synthesizing ascorbic acid, which is an antioxidant and cell reductant, and the improvement of antioxidant capacity is one of the characteristics of plant resistance [20]. Galactinol is also involved in inducing resistance in plant systems [21]. Glyceric acid is up-regulated in Rhizoctonia solani-infected resistant rice and participates in photorespiration and plant defense response regulation [9]. Thus, this indicates that the above illustrated organic oxygen compounds are unique metabolites accumulated in infected transgenic rice, and their biochemical and structural defense state with high antioxidant capacity were at the activation stage. The amino acid and carbohydrate metabolism pathways to which these accumulated metabolites belong were also activated. Therefore, the resistance in infected transgenic rice was strengthened.
Linoleic acid and linolenic acid are lipids that were found to accumulate at 72 h. Linolenic acid is a precursor of jasmonic acid [10] that plays a vital role in plant defense response. Thus up-regulated linolenic acid is a unique metabolite of infected transgenic rice. Subsequently, a further experiment validated an increase in jasmonic acid concentration at 120 h. Consistently, linoleic acid involved linoleic acid metabolism at 72 h.
L-glutamine is involved in stress response in plants [22]. Kan et al found that glutamine can be used in the act of a signal molecule to modify expression patterns of plants. The prompt evocation of transcription factors indicates that glutamine can effectively intensify its signaling and engage with diverse signaling communication pathways to determine the growth of plant and its stress responses. Therefore, glutamine is an underlying effective amino acid that takes a critical role in nutrition and signaling transductions of plants [22]. L-tryptophan was found to accumulate at 72 h, indicating that the infected transgenic rice continues to grow. Oxalate has a dual role by inhibiting plant defense responses at the early stages of infection while also inducing reactive oxygen species and cell death at later stages of infection [23]. In our study, the concentration of oxalate decreased at 72 h, which was beneficial as it enhanced the ability of ROS clearance in infected transgenic rice and inhibited cell death. Most of these unique metabolites are involved in the citrate cycle (TCA cycle), thus indicating the activation of this pathway.
Benzenoids accumulated at different time-points in M. oryzae-infected transgenic rice, in which salicylic acid and anthranilic acid (Vitamin L1) accumulated at three-time points. Anthranilic acid (AA) is a vital precursor in the synthesis of major auxin, indole-3-acetic acid (IAA) [24], essential for maintaining the growth of infected transgenic rice. While salicylic acid plays a vital role in defense response [25], suggesting that MoSDT1 may play a role in balancing rice growth and defenses. Dopamine has an anti-aging effect that regulates the absorption and transport of nutrients by plants, which ultimately affects the overall growth of plants [26]. Tyramine accumulation, which is beneficial to increase resistance, was induced when barley was infected by Bipolaris sorokiniana [27]. Therefore, the accumulation of dopamine and tyramine in transgenic rice is beneficial to improve resistance. The metabolic pathways involved with tyrosine, tryptophan, and phenylalanine metabolism were thus activated.
Unclassified metabolites, such as vitexin, riboflavin, sinapyl alcohol, diosmetin, myo-inositol, D-biotin, and camptothecin, and alpha-pinene were found to accumulate at varying degrees at different time points. Both vitexin and apigenin have antiviral, antioxidant, and anti-cancer effects in plants [28]. L-tyrosine, an aromatic amino acid, is a precursor of lignin in plants of the grass family [29]. D-biotin plays an essential role in combating M. oryzae infection and promoting rice growth [30]. Meanwhile, D-biotin could also inhibit ROS accumulation and the appearance of lesion-mimic [30]. Sinapyl alcohols are products of lignin monomer synthesis [11]. Diosmetin, a flavonoid compound with antioxidant activity, protects plant tissues from oxidation [25]. Myo-inositol also plays a crucial role in the tolerance of abiotic stress [31]. These results indicate that these metabolites play an essential role in maintaining the growth of Magnaporthe oryzae-infected transgenic rice, keeping a dynamic balance of ROS accumulation and clearance, initiating structural defense response, and the restriction of spotted-lesion expansion and enhanced resistance of MoSDT1-transgenic rice.
Collectively, during the infection of MoSDT1-transgenic rice with M. oryzae at three-time points, organic oxygen compounds, lipids, and lipid-like molecules, benzenoids, organic acids, and derivatives, together with many unclassified primary defensive metabolites accumulated. Metabolic pathways involved with these metabolites are attributed to carbohydrate and amino acid metabolic pathways, which play a key role in stress response [19]. Therefore, these activated pathways participated in rice defense responses. This study found that at 72 h, the accumulation of defensive metabolites reached a peak, indicating that 72 h was the time for most active defense response, which was also the time point at which brown spotted lesions began to appear [32]. The increase in the number of accumulated defensive metabolites at this time is conducive to strengthen the defense response of MoSDT1-transgenic rice, restrict the spread of spotted lesions and increase the resistance of rice.
The accumulated metabolites identified in this study, such as diosmetin, D-biotin, tyramine, galactinol, alpha-Pinene, L-tryptophan, linolenic acid, sucrose, raffinose, fructose, and L-glutamine, can serve as core defensive metabolites for strengthening the resistance of rice. Hence, the predominantly enriched carbohydrate and amino acid metabolic pathways are the keys to the defense response.
Crosstalk between multiple defensive metabolite response genes and plant hormones in M. oryzae-infected MoSDT1-transgenic rice
Linolenic acid and linoleic acid are precursors of jasmonic acid synthesis [10]. Salicylic acid and jasmonic acid are important hormones of plants, which take crucial roles in defense response and stable growth of rice [33]. Sun et al examined changes in the phosphorylated proteomics of rice response to SA, suggesting that SA-mediated phosphorylation regulation may lead to different resistances of the two rice varieties [34]. OsVQ13 positively regulates JA signal by activating OsMPK6-OsWRKY45 signaling pathway in rice [35]. JA reduces sensitivity of RBSDV infection by inhibiting the brassinosteroid (BR) pathway [36]. However, it is not clear about the JA/SA that contribute to defense response in rice.
In this study, these two metabolites and salicylic acid were identified to accumulate in M. oryzae-infected MoSDT1-transgenic rice at 72 h. Meanwhile, the contents of JA, JA-Ile, and salicylic acid were higher in infected transgenic rice than infected wild-type rice.
In this study, JA signaling pathway genes (OsCOI1b, OsJAZ1, OsJAZ9, and OsMYC2), JA early response genes (JiOsPR10 and OsbHLH35) and defense-related genes (OsPR4a and OsPR10a) were inhibited at 72 h and 120 h. Further analysis of JA synthesis pathway-related genes revealed that OsLOX1/3, OsHPL3, OsOPR1, OsOPR7, and OsJMT1 were up-regulated at 72 h. In addition, OsLOX3 accumulated linolenic acid at 72 h, and subsequently, the JA amount reached a peak at 120 h. It thus indicates that OsLOX3 was mainly involved in converting linolenic acid to 13-hydroperoxide. OsLOX1 and OsLOX3 are similar in genetic structure and both are involved in pathogen defense responses [37], OsHPL3 mediates SA and JA increase and enhances rice defense response [36, 38]. Increased JA and SA levels were noticed in this study, demonstrating that OsHPL3 regulates JA and SA content in response to pathogen infection. In addition to its role in JA synthesis, OsJMT1 is also primarily involved in responding to pathogen infection, during which OsOPR7 got involved in JA biosynthesis in plants. However, OsOPR1 plays a role in the rice self-defense response [39], indicating that the jasmonic acid synthetic pathway is activated, and genes in this pathway are involved in both jasmonic acid synthesis and defense response. It is, therefore, speculated that MoSDT1 might regulate jasmonic acid synthesis pathway-related genes to function in jasmonic acid synthesis and defense responses.
On the one hand, the up-regulation of OsEDS1/OsPAD4 promotes SA biosynthesis, while on the other, it maintains SA-related resistant responses, strengthening the plant's internal immune system [40]. It was reported that knock-out of rrsRLK gene can induce significant up-regulation of OsPR1a, OsPR1b, OsLOX and RBBTI4 [41]. Thus indicating that the SA signaling pathway is activated, and the internal immune system is enhanced when transgenic rice is infected.
The expression level of the OsOXO4 gene was significantly up-regulated in M. oryzae-infected MoSDT1-transgenic rice at 72 h. Meanwhile, the concentration of oxalate in infected transgenic rice was decreased, indicating that OsOXO4 plays a role in scavenging oxalate at 72 h, avoiding the accumulation of reactive oxygen species and inhibition of cell death at the late stage of infection in transgenic rice. The expression level of raffinose synthase gene Os01g0170000 and galactitol synthase gene OsGolS1 in M. oryzae-infected MoSDT1-transgenic rice was higher than the wild type. Salicylic acid (SA) and nitric oxide (NO) also play a role in protecting plants from oxidative damage [42]; thus, the oxidative damage of infected transgenic rice is lower than the wild type rice. Therefore, OsOXO4, Os01g0170000, and OsGolS1 play a role in protecting M. oryzae-infected MoSDT1-transgenic rice from infection with reactive oxygen species and inhibit drastic cell death at the late stage of the infection.
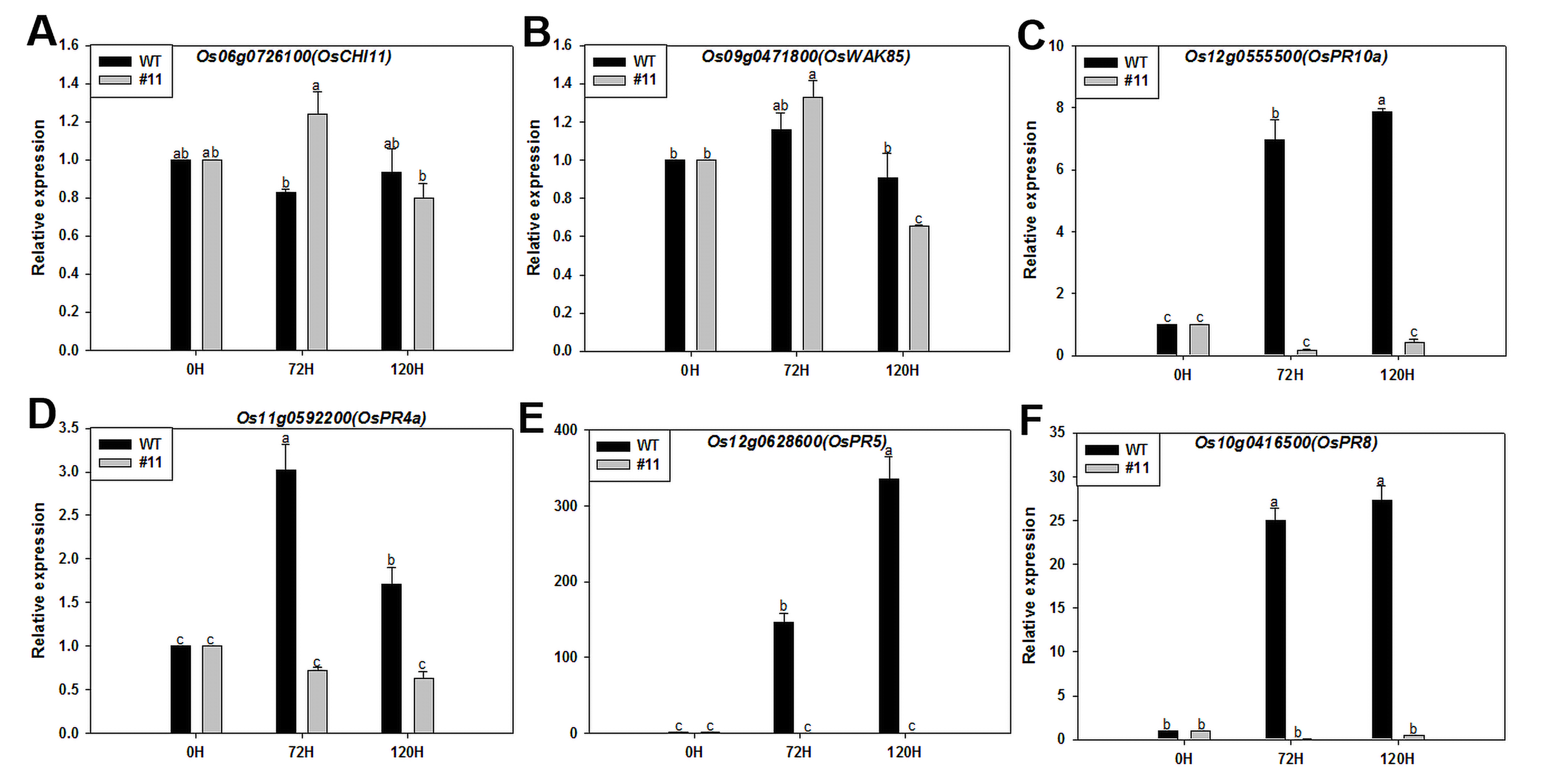
Chitinase and cell wall-associated kinases play a major role in plant defense and resistance to pathogenic fungal infection [43]. Cellulose synthase complex is regulated by metabolic signals related to plant carbon status [44]. Most of the fructose entering the cytoplasm, after phosphorylation by fructokinase (FRK), become components of cell wall synthesis functioning in xylem vessels, fibers, and overall vasculature development [45]. In this study, the expression levels of cell wall-associated kinase genes OsCHI11 and OsWAK85, sucrose synthase gene OsRSUS1 and fructose synthase gene OsFRK–2 in MoSDT1-transgenic rice were significantly higher at late stages of the infection compared to the wild-type rice. Thus, indicating that these up-regulated genes are involved in cell wall remodeling or construction to enhance the physical barrier in MoSDT1-transgenic rice.
To summarize, SA, JA, raffinose, fructose, and sucrose accumulated in M. oryzae-infected MoSDT1-transgenic rice. Meanwhile, JA synthesis, SA synthesis/receptor gene, raffinose, fructose, and sucrose synthesis enzyme genes, and the cell wall-associated kinase gene were up-regulated. In contrast, genes involved in the JA signal gene were suppressed. These results indicate that JA and SA play a synergistic role in defense responses in MoSDT1-transgenic rice. ROS clearance-associated gene (OsOXO4) and raffinose synthesis genes (Os01g0170000, OsGolS1) play a role in protecting active oxygen damage. Moreover, the Fructose/sucrose synthase gene and cell wall-associated kinase genes are essential in cell wall remodeling.
Improved resistance to M. oryzae by exogenous treatment with defensive metabolites
Studies have shown that biotin, galactitol, citrate, tyramine, raffinose, L-glutamine, L-tryptophan, dopamine, galactitol, terpinene, L-tryptophan and other metabolites serve as defensive signals in plant defense system [22, 46]. Rice infected with M. oryzae develop brown spotted lesions on leaves at 72 h and enter the time-point of dead body nutrition stage [32]. This is the time-point when metabolite accumulation is detected in infected transgenic rice, and most of these metabolites are associated with pathogen resistance. In addition, studies have also shown that exogenous application of metabolites (such as GABA, dopamine, proline, tyramine) can increase plant resistance [26, 47-49]. From the metabolites accumulated in MoSDT1-transgenic rice infected by M. oryzae, six metabolites (galactinol, tyramine, L-glutamine, L-tryptophan, pinene and dopamine) were selected to treat M. oryzae-infected rice in two ways exogenously. Among these six metabolites, the treatment significantly increased rice resistance at 72 h, and its low concentration also played a significant role in improving rice resistance. Therefore, low-concentration metabolites as defensive weapons have the most apparent control response when brown spotted lesions of rice blast begin to form (72 h); however, the underlying mechanism involved requires further investigation.
To summarize, among the defensive metabolites accumulated at different stages of the pathogen-infected plant, six defensive metabolites were selected to treat the plants exogenously at a specific time-point of the infection to improve plant resistance. Thus this makes exogenous application of defensive metabolites an environmental control strategy for future applications.
{kind=link}