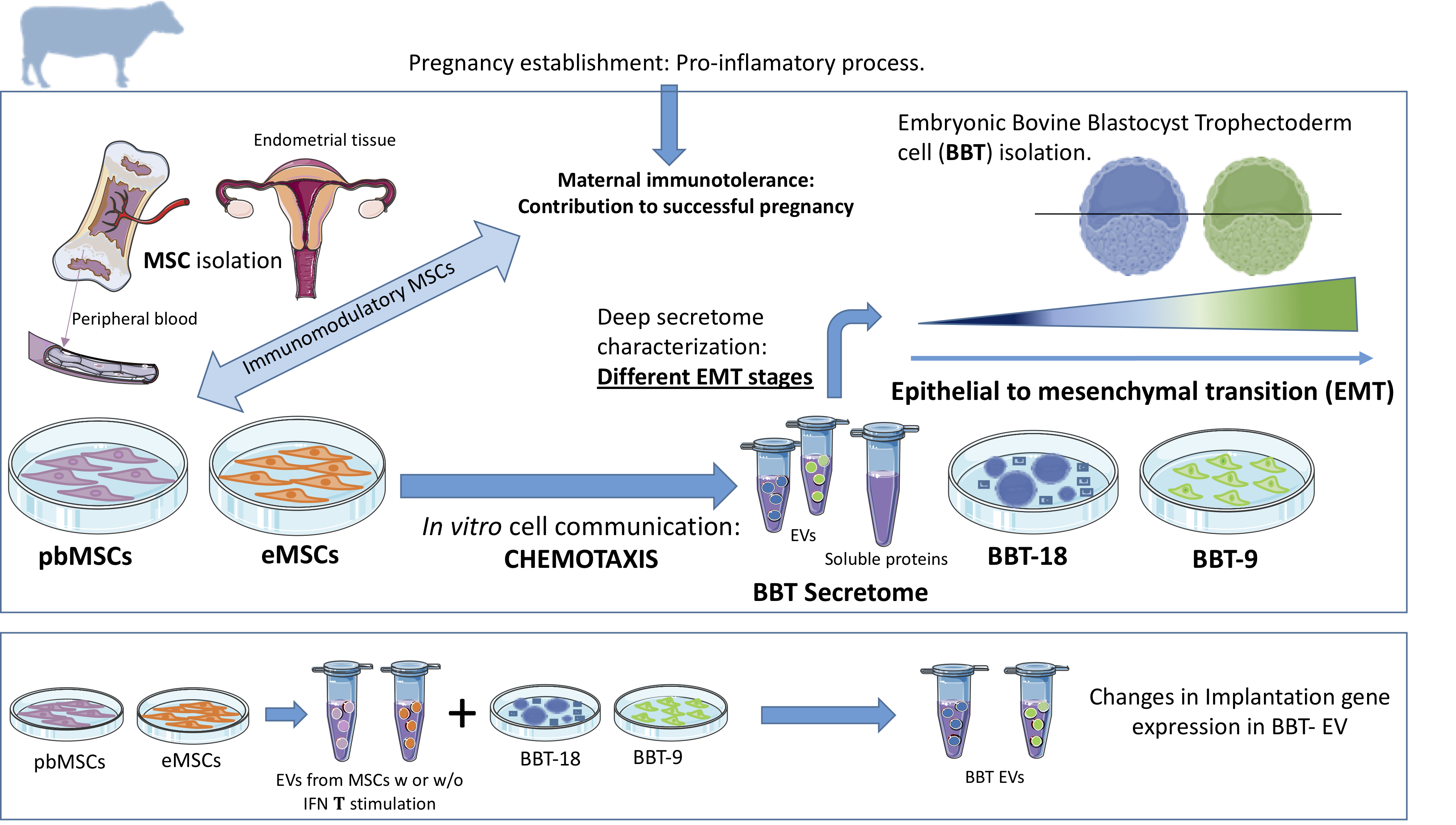
Recently, our group characterized for the first time endometrial mesenchymal stem cell lines (eMSC) [9]as well as peripheral blood MSC (pbMSCs) lines [8]. While eMSC showed a reduced migratory capacity in response to the implantation cytokine IFN-τ [9], pbMSC showed chemotactic behavior to both inflammation (TNFα, IL1β), embryo implantation stimuli (IFN-τ) or BBT-secretome, suggesting that the embryo secretome plays a role in ensuring the retention of eMSC and the active recruitment of MSCs from bone marrow during early pregnancy to repress the immune response to prevent the embryo rejection by the maternal organism [8].
During bovine pregnancy, trophoblast adhesion and placental formation has been reported to require a gradual loss of epithelial characteristics but without the acquisition of full mesenchymal characteristics. [44]. This partial EMT concurs with gene expression changes associated with EMT in the bovine trophectoderm following conceptus attachment to the luminal epithelium [44]. After conceptus implantation the trophectoderm looses the adherents junction molecule CDH1, and gained the expression of mesenchymal markers, such as VIM and CDH2, maintaining however, the expression of the epithelial marker cytokeratin. Trophectoderm EMT was thereafter shown to be regulated by the endometrium via activin A and FLST1 on days 20–22 [21]. Trophoblast binucleate cells in the bovine placentome showed a cytoplasmic distribution of CH1 and b-catenin translocation into the nucleus [28], suggesting a role for CDH1–b-catenin axis in trophoblast differentiation. On day 22 trophoblasts CDH2 is highly expressed, so that an increment in CDH1 degradation could also be involved in the further reduction of its expression. Hence, the loss of CDH1 as the conceptus attaches to luminal epithelium, may play a crucial role in gene expression transition required for the successful progression from implantation to placentation. Regarding our BBT primary cultures, we observed that the expression of CDH1 was overrepresented in BBT-18 soluble fraction but not in BBT-9, while BBT-9-secretome is expressing VIM while maintaining cytokeratin expression. These parameters would reflect the two above mentioned phenotypes; epithelial and mesenchymal, respectively. CDH2 was expressed in vitro when trophectoderm cells were co-cultured with endometrial epithelial cells [18], but has not been reported in monocultures of trophectoderm cell lines possibly explaining the lack of detection of CDH2 in the BBT-secretome.
EMT induction is regulated at the molecular level by a variety of growth factor signals, in particular transforming growth factor β (TGF-β), hepatocyte growth factor (HGF), epidermal growth factor (EGF), fibroblast growth factor (FGF), Wnt proteins, and IL-6 [35]. TGF-β is a multifunctional cytokine that is considered the main inducer of EMT. The TGF-β signaling pathway plays an important role in regulating cell proliferation, differentiation, invasion, migration, apoptosis, and microenvironmental modification, and stimulates pathophysiological EMT and metastasis [29]. TGF-β and FGFR1 proteins were found to be overrepresented in BBT-9-secretome, suggesting that these proteins could be responsible for the regulation of EMT in the bovine trophectoderm. In addition, it has been reported that TGF-β is a soluble factor produced by MSCs, which mediates the suppression of T-cell proliferation [13]. Therefore, the BBT-9, thanks to its mesenchymal phenotype, could play an immunomodulatory role via EVs signaling, aimed at guaranteeing the survival of the embryo. Our group has also reported the immunomodulatory capacity of eMSC and pbMSC by inhibiting the proliferation of human T lymphocytes [8, 9].
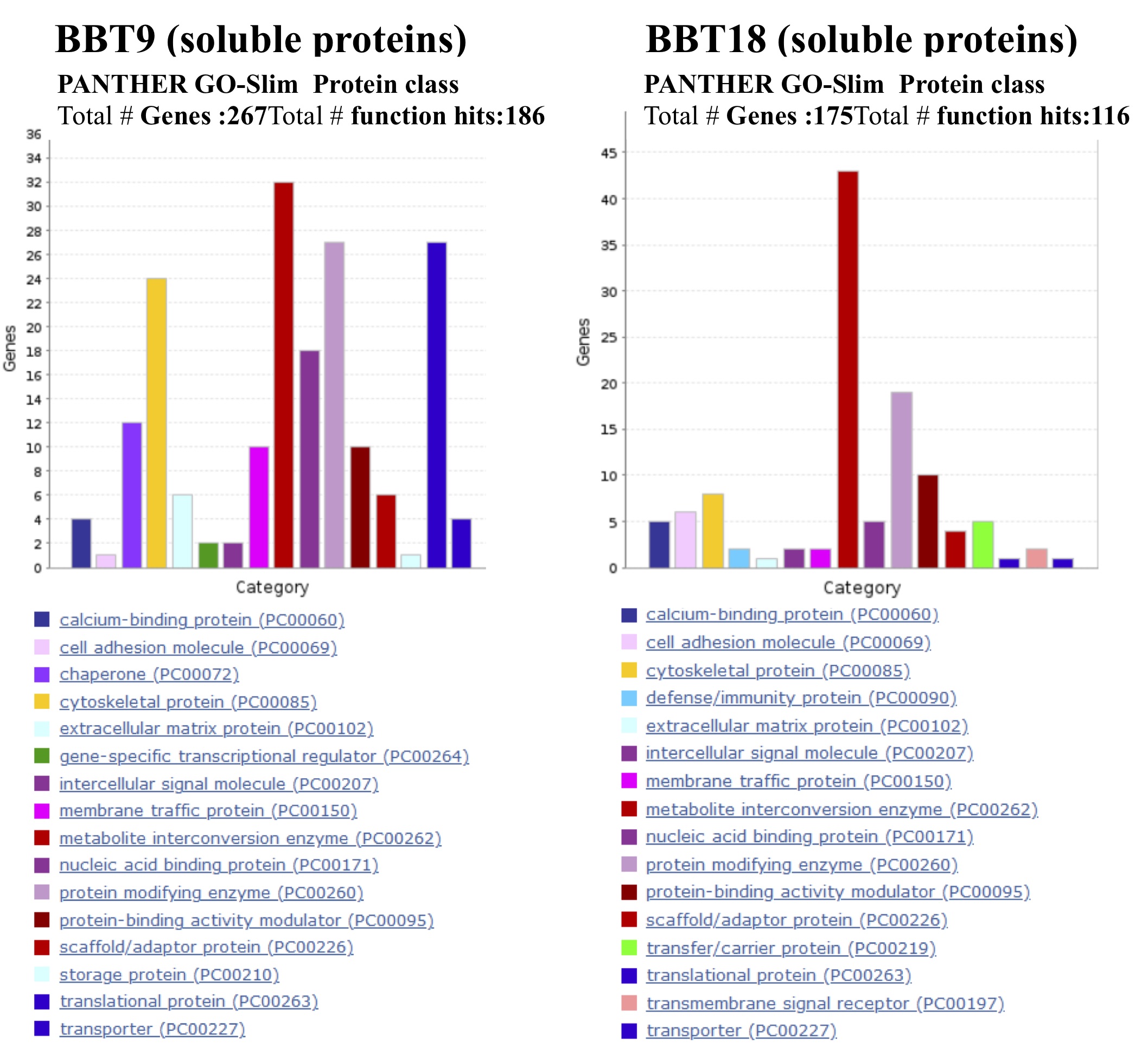
During EMT, FGF increases the expression of vimentin and induces the activity of MMP2, increasing cell mobility. FGF also causes changes in the actin cytoskeleton allowing anchorage-independent growth [33]. Vimentin and MMP2 are overrepresented in BBT-9-EVs and BBT-9-soluble fractions respectively. Although bovine trophoblasts do not penetrate the endometrium, upregulation of MMP2 metalloproteinase suggests that it could play a role in the non-invasive trophectoderm. FGF1, the ligand for FGFR1, overrepresented in BBT-9-soluble fraction, is also known to upregulate MMP13, resulting in EMT induction [6].
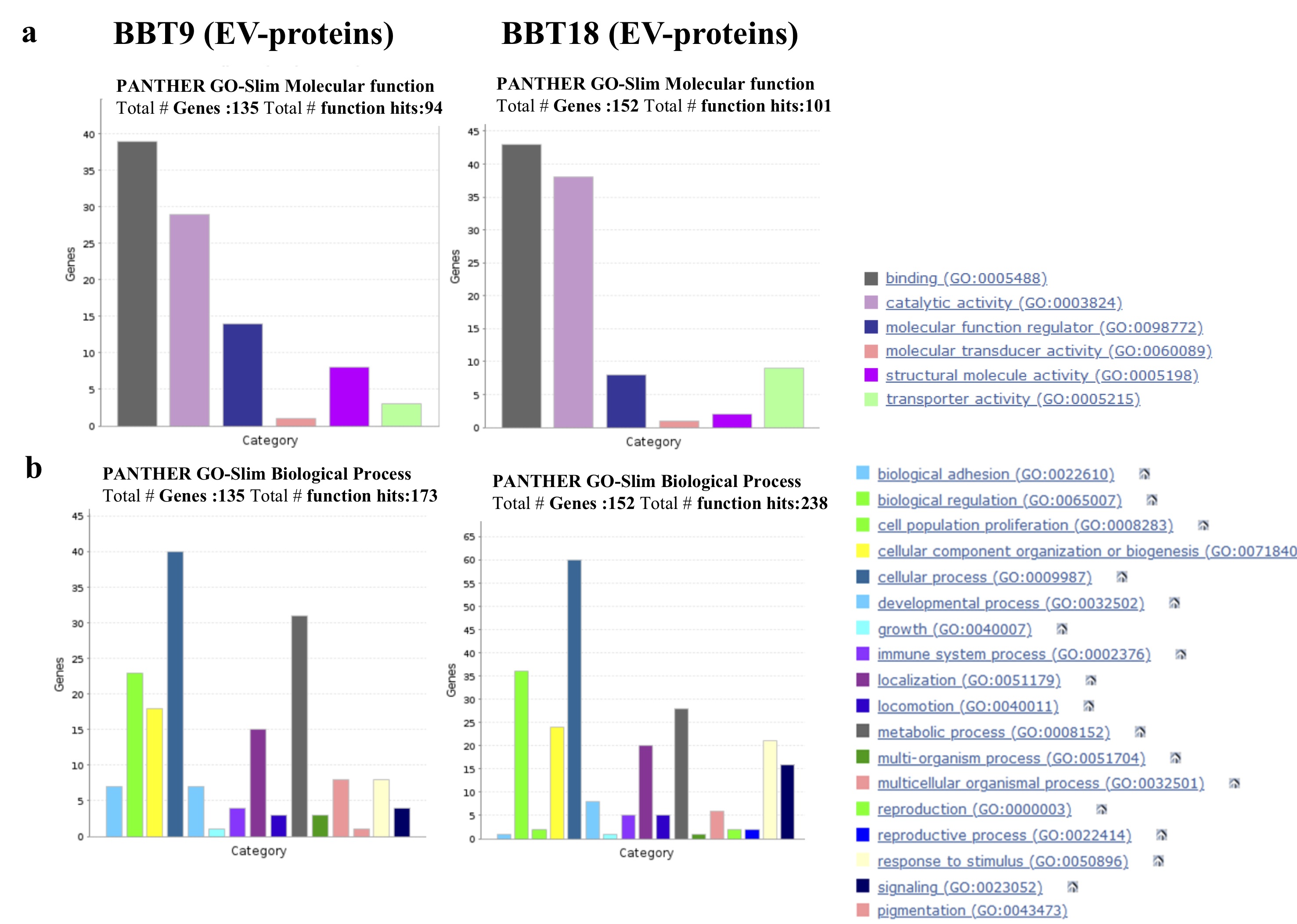
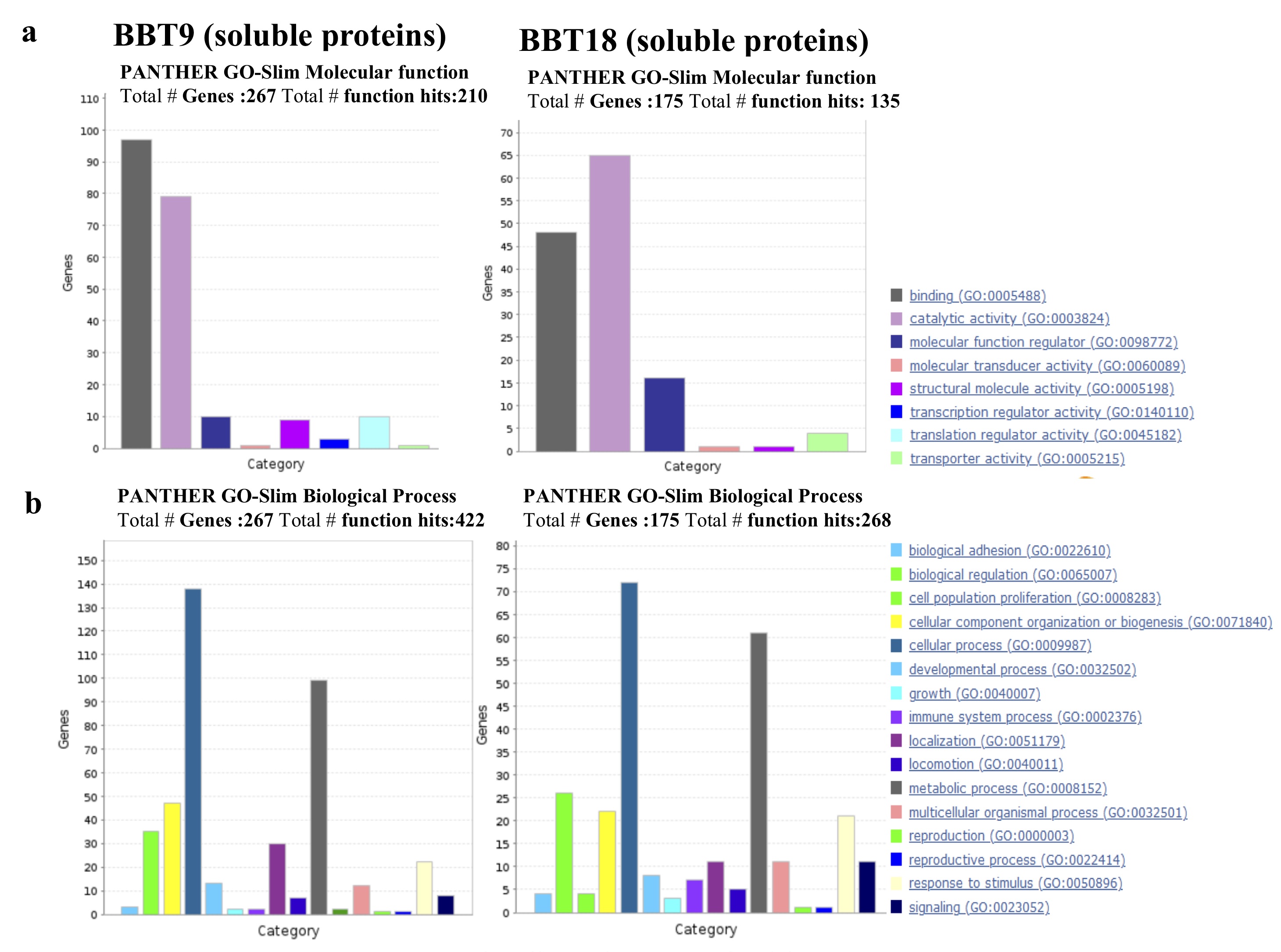
Other characteristic EMT markers such as the transcription factors, SNAI2, ZEB1, ZEB2, TWIST1, TWIST2, and KLF8S were not found in EVs or soluble proteins from BBT-secretome as we could expect since they are nuclear proteins. Likewise, we were also unable to detect pluripotency markers (OCT4/POU5F1, SOX, NANOG) or characteristic markers of trophectoderm (ELF5, EOMES, GATA5) in EVs or soluble proteins from BBT-cargo, although some of the mentioned markers could be detected by RT-PCR in our BBTs [32].
After EMT, a micro-angiogenesis process related to the uterine vascularization is necessary for adequate implantation [16] and placenta formation. We were able to detect eleven proteins involved in angiogenesis pathways in the BBT secretome, from which six participate in the vascular endothelial growth factor (VEGF) pathway. The VEGF signaling pathway is essential for all stages and processes involved in vascular development (vasculogenesis, angiogenesis, and lymphangiogenesis). VEGF is the main factor that regulates angiogenesis in bovine pregnancy. Interestingly, BBT-9 secretome showed an overrepresentation of several angiogenic related factors: Fibroblast growth factor receptor 1 (FGFR1), RHO GTPase-activating protein 1 (ARHGAP1), RHO-related GTP-binding protein (RHOC), and Vascular cell-adhesion molecule (VCAM1). However, in BBT-18 secretome only the Serine/threonine-protein kinase A-RAF (ARAF) and Heat Shock Protein Beta-1(HSPB1) from this pathway, were found to be overrepresented.
In sum, both the overexpression of EMT markers and the higher detection of angiogenic factors in the BBT-9 protein profile would support the notion that BBT-9 is exhibiting a more advanced developmental stage near to an EMT stage. However, we could not detect in the secretome of BBT the typical bovine pregnancy angiogenic related markers such as VEGF family proteins or its receptor, Angiopoietin (ANGPT)-2/ANGPT-1 [16, 17] due likely to an early developmental stage.
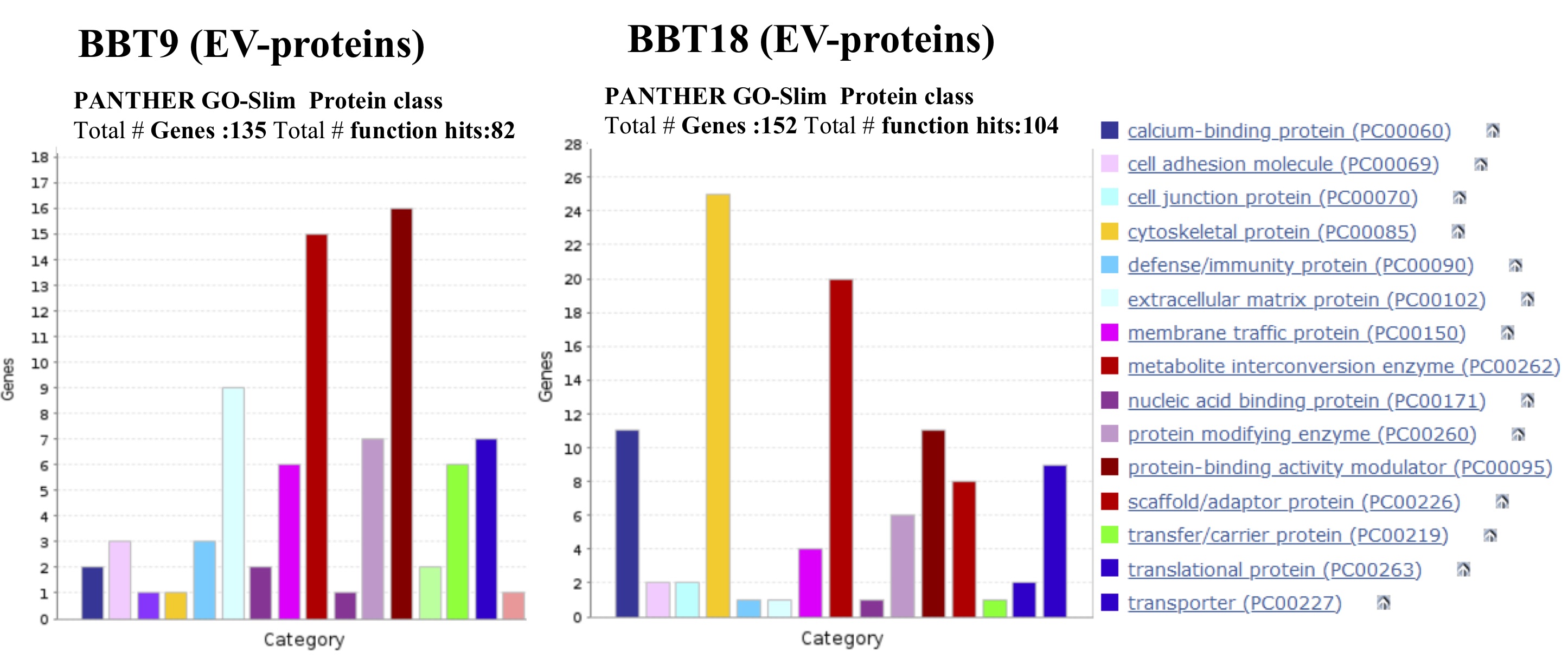
In the bovine species, the expression of integrins (ITGs) has been characterized at the uteroplacental interface during the periods of trophectoderm attachment and placentation [30]. ITGαV (overrepresented in BBT-9-EVs cargo), in combination with the b5 subunit is known to bind to Osteopontin (SPP1) [10]. ITGβ1 is also overexpressed in BBT-9-EVs cargo. It can form heterodimers with the α4 chain, also known as very late antigen-4 (VLA4) often detected in mesenchymal stem cells [3], and α8 subunit resulting in alternative receptors for osteopontin SPP1 [34]. Yamakosi et al observed that the subunits of SPP1-binding ITGs are upregulated during the embryo attachment process and indicate their possible involvement in the trophoblast adhesion to the endometrial epithelium in cows [44]. Moreover, ITG on EVs was reported to dictate organ-specific uptake of EVs to initiate pre-metastatic niche formation in a tumor scenario [44]. Analogously, integrin expression profiles of trophectoderm-secreted exosomes could be relevant to direct maternal MSCs to the implantation niche and could be used as prognostic factors to predict good maternal immunoregulation to avoid embryo rejection.
As we have described previously, a correct balance that coordinates active immunity and tolerance during the contact between mother and conceptus is critical. Moreover, it is known that MSCs from the placenta or decidua are involved in the induction of this maternal immune tolerance [23, 43]. In mice, MSCs from bone marrow are involved in the reduction of the embryo resorption rate by regulating the function and phenotype of macrophages and T cells at the maternal-fetal interface [26]. ITG α4β1 major counter ligand, vascular cell adhesion molecule-1 (VCAM-1), plays an important role in leukocyte recruitment during an immune response [5]. Bai et al reported that uterine VCAM-1 expression was minimal in day 17 cyclic and pregnant animals, but increased between days 20 and 22 of pregnancy [3]. The authors also reported that VCAM-1 expression in CT-1 cells (a trophoblast primary culture cultured onto STO mouse feeder cells [44] was up-regulated with the use of uterine flushings. VCAM-1 is found to be overrepresented in BBT-9-EVs cargo. Galectin 3 (LGALS3), also play central roles in immune system regulation, shaping both innate and adaptive responses in physiological and pathological processes [25]. Galectin 3, which is overrepresented in BBT-9-EVs cargo, is first expressed in the trophectoderm cells of the implanting embryo and has been implicated in the process of implantation [12].
We have demonstrated that both eMSC and pbMSCs are capable to migrate in response to BBT chemotactic stimuli reinforcing the idea of their participation at the pregnancy establishment. Considering that BBT-9 shows developmental features nearer to EMT than BBT-18, it would suggest that it represents a more advanced developmental stage, closer to embryo implantation than BBT-18. Therefore, our chemotactic data would support that epithelial embryonic trophectoderm (BBT-18) stimulates chemotactic migration of maternal MSCs from the endometrium through soluble and EVs mediators, while it attracts peripheral MSC only through soluble mediators. In contrast, when the embryonic trophectoderm already presents mesenchymal characteristics (BBT-9), it stimulates the migration of endometrial or stimulates peripheral maternal MSCs through both soluble and EVs-cargo proteins, traveling long distances and at high speed. Therefore, at late implantation stages (BBT-9) secretome-dependent signaling could provoke a highly intensified call-effect in MSCs to ensure embryo implantation in that critical point of pregnancy.
We next analyzed the changes in the secretome of embryonic trophectoderm as a result of the uptake of maternal MSC-EVs or treatment with endometrial cytokines present in the uterine fluid in the peri-implantation period. Activin A is a known member of the TGF-ß superfamily and FSLT1 is an inhibitor of activin A [19]. Kusama et al reported that FLST1 increased on day 20 uterine flushing and decreased on day 22, whereas they found an elevated activin A on day 20 and further increased on day 22. [21]. Kusama et al reported Activin A-induced EMT marker expressions were inhibited by FLST in the embryonic trophectoderm cells CT-1 [21]. To analyze the interaction between activin A and FLST1 in bovine trophoblasts with epithelial or mesenchymal phenotype, BBT-9 or BBT-18 cells were treated with Activin A or Activin A + FLST1.
As a readout we decided to focus on the possible variations in the secretion pattern of a selected group of proteins related to the process of invasive growth and thus potentially involved in the regulation of embryo implantation: TDGF1, HSPH1, MMP2, and PEG3 [11, 14, 27, 46]. Our embryonic trophectoderm primary cultures either with epithelial or mesenchymal phenotype, responded to Activin A or Activin + FLST1 stimulations by secreting the implantation proteins in EVs-cargo. Mitko et al. reported a marked increase in the expression of TDGF1 at day 12, connected with the process of invasive growth and regulating the embryo implantation [27]. Hatayama et al reported a marked increase in the expression of HSPH1 in mouse embryos between days 9 and 12, coinciding with organogenesis, and they attributed to HSPH1 a relevant function in organogenesis during embryonic development [15]. Later, Yuan et al also reported the presence of HSPH1 in rat embryos [45]. In our in vitro model of communication between maternal MSCs and embryonic trophectoderm through EVs, we observed a marked expression of HSPH1 in EVs from trophectoderm with the epithelial phenotype (BBT-18), that was further increased in the presence of Activin A. In contrast, the expression of HSPH1 in EVs from embryonic trophectoderm with mesenchymal phenotype was much lower. MMP2, a member of the matrix metallopeptidase family, which is involved in the breakdown of extracellular matrix in normal physiological processes, and PEG3 are up-regulated on Day 13 in vivo conceptuses [44].
In conclusion, we have confirmed different epithelial-mesenchymal transition stages in embryonic trophectoderm primary cultures. The embryonic trophectoderm secretome contains early pregnancy-related proteins and angiogenic markers both as cargo in EVs and the soluble fraction. We have demonstrated the functional transfer of protein-containing secretome between embryonic trophectoderm and maternal MSC and its chemotaxis capacity, thus suggesting that this system could be used as an in vitro model of cell-to-cell communication through EVs and soluble proteins. In a complementary way, the pattern of secretion of implantation proteins in trophectoderm-EVs changes depending on: i) its epithelial or mesenchymal phenotype; ii) as a result of the uptake of eMSC- or pbMSC-EVs previously stimulated or not with embryonic signals; and iii) because of stimulation with endometrial cytokines present in the uterine fluid in the peri-implantation period.
Figure 7 shows a chronologic schematic representation of BBT identified proteins associated with the main reproductive or EMT events.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}