Motivated by a basal activity of wild-type CtXR on rac-2-phenylpropanal (kcat/Km,rac 130 s− 1M− 1, kcat 0.05 s− 1), we tested a number of substrate binding mutants for activity and enantioselectivity in the reduction of 2-phenylpropanal. The used enzyme is a member of the aldo-keto reductase superfamily (AKRs) and converts xylose to xylitol in the central sugar metabolism of its native host Candida tenuis. CtXR shows, like many other AKRs, broad substrate specificity. Its biocatalytic applicability was, however, limited by moderate catalytic activity and low stability of the wild-type enzyme. Several substrate-binding site mutants with improved specificity for aromatic ketones were used in the synthesis of (R)-ethyl mandelates and (S)-phenylethanols (13, 14).
Ct XR mutants
The substrate-binding cavity of aldo-keto reductases is mainly formed by residues from three large and flexible loops (15, 16). Loop flexibilities provide the structural basis for relaxed substrate specificities but complicate rational engineering (17). A binding mode of the natural substrate D-xylose (open chain form) was previously modelled with C-1 of xylose within hydride-transfer distance above the nicotinamide C-4 and the carbonyl oxygen hydrogen-bonded to the general acid catalyst Tyr-52. Therein, the aldehyde hydrogen pointed towards the indole ring of Trp-24, the C2 hydroxyl interacted with Asn-310 and C3, C4 and C5 hydroxyls with Asp-51 (18). Here, we probed CtXR wild-type and single-point mutants of the main substrate recognition residues Trp-24, Asn-310 and Asp-51 as catalysts for 2-phenylpropanal reduction. The replacement of Trp-24 by smaller phenylalanine and tyrosine increased activities on bulky ketone-substrates (13, 19). Asp-51 contributes the most to the relative polarity of the binding site; substitution by alanine led to improved selectivity for the aromatic ketone o-chloroacetophenone (14).
Kinetic constants of 2-phenylpropanal reduction by CtXR variants
Table 1 summarizes results of a steady-state kinetic analysis of NADH-dependent reduction of racemic and (S)-2-phenylpropanal by CtXR wild-type and mutants.
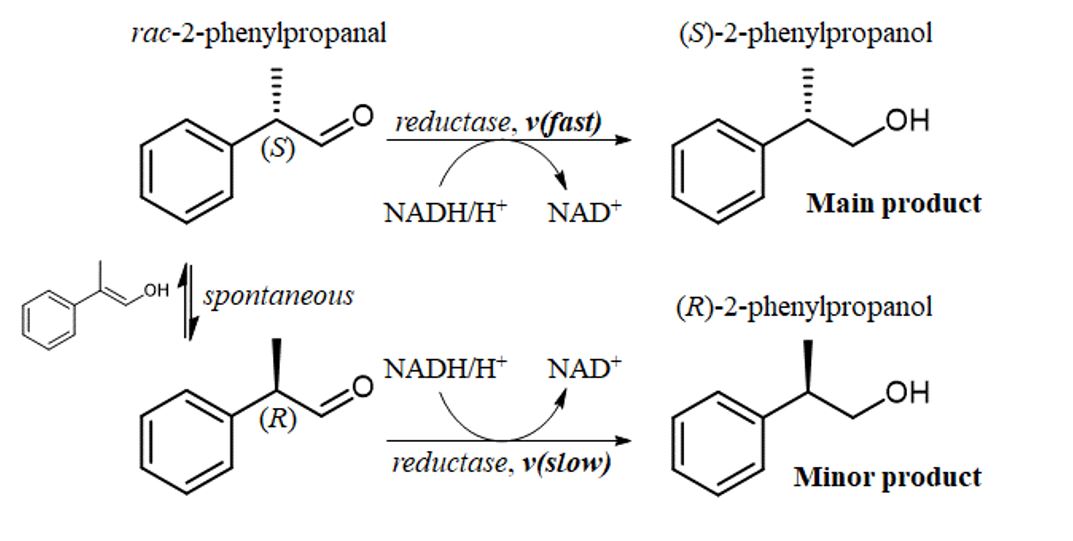
Racemic substrate. The wild-type enzyme showed a specificity constant (kcat/Km,rac) of 130 s− 1M− 1 reflected by a Km,rac value of 350 µM and a kcat value of 0.05 s− 1. The mutants W24F and W24Y displayed only 8 to 10 % of the wild-type activity. While the N310A mutant had a specificity constant similar to the wild-type, the N310D mutant showed no activity. CtXR D51A stood out with a kcat/Km,rac of 28·103 s− 1M− 1 composed of a Km,rac value of 170 µM and a kcat value of 4.8 s− 1. Replacement of the charged aspartic acid by alanine led to 215-fold higher catalytic efficiency compared to the wild-type. Likewise, introduction of an additional aspartic acid in the substrate binding pocket (N310D mutant) abolished the activity with 2-phenylpropanal. Note that used CtXR mutants showed 35 to 100-fold reduced catalytic activity towards the natural substrate xylose (19). Enlargement of the substrate binding pocket by replacement of the bulky Trp24 decreased the enzyme's activity towards 2-phenylpropanal.
(S)-2-phenylpropanal. Kinetic parameters obtained with the racemic substrate and the (S)-2-phenylpropanal were compared and the ratio of the specificity constants formed (kcat/Km,S)/(kcat/Km,rac) (Table 1). For the extreme case of sole activity with the S-enantiomer, a ratio of ~ 2, for equal acceptance of S- and R-enantiomer a ratio of 1 and for preference of the R-enantiomer ratios < 1 were expected. The wild-type showed a ratio of 1.23 and preference for the S-enantiomer. The D51A mutant showed a ratio of 1.54 and hence a stronger preference for the S-enantiomer. The W24F, W24Y and N310A mutants, however, displayed ratios < 1 and therefore preference for the R-enantiomer.
Table 1
Apparent kinetic parameters of CtXR wild type and mutants for NADH-dependent reduction of racemic and (S)-2-phenylpropanal.
| |
rac-2-phenylpropanal
|
(S)-2-phenylpropanal
|
Ratio
specificity constants
|
|
CtXR
|
kcat/Km,rac (s-1M-1)
|
Km,rac (µM)
|
kcat/Km,S (s-1M-1)
|
Km,S (µM)
|
(S)-aldehyde /
racemic aldehyde
|
|
wild-type
|
130
|
350
|
160
|
450
|
1.23
|
|
D51A
|
28·103
|
170
|
43·103
|
120
|
1.54
|
|
W24F
|
13a
|
n.a.
|
12
|
n.a.
|
0.92
|
|
W24Y
|
10a
|
n.a.
|
9
|
n.a.
|
0.90
|
|
N310A
|
88
|
280
|
68
|
330
|
0.77
|
|
N310D
|
no activity
|
no activity
|
no activity
|
no activity
|
-
|
| aLimited substrate solubility prevented saturation of the enzyme; kcat/Km was calculated from the slope of the Michaelis–Menten plot where the rate is linearly dependent on the substrate concentration. |
Reduction of rac-2-phenylpropanal by isolated D51A CtXR
We used isolated CtXR D51A in bioreductions of 0.5 mM rac-2-phenylpropanal. The substrate (logP 2.11, https://scifinder.cas.org/) displayed a maximal solubility of 0.5 mM in buffer. Product concentrations and e.e. values obtained at enzyme concentrations between 240 and 0.6 U/mL are listed in Table 2. As expected, the product e.e. values increased with decreasing amounts of enzyme. Unexpectedly, reactions stopped after 2 h and maximal product concentrations of 120 µM were achieved (for a time course see the Supplementary data). We suspected enzyme deactivation as cause of low conversions and used a whole-cell biocatalyst based on D51A CtXR in further experiments.
Table 2
Conversions and product e.e.-values of rac-2-phenylpropanal reduction by isolated D51A CtXR. a
|
CtXR (U/mL)b
|
(S)-Phenylpropanol (µM)
|
e.e. (S)-Phenylpropanol
(%)
|
|
D51A (240)
|
25
|
23
|
|
D51A (20)
|
16
|
38
|
|
D51A (3.4)
|
11
|
≥ 99
|
|
D51A (0.6)
|
4
|
≥ 99
|
| aReaction time 2 h. bMeasured with 0.5 mM rac-2-phenylpropanal. |
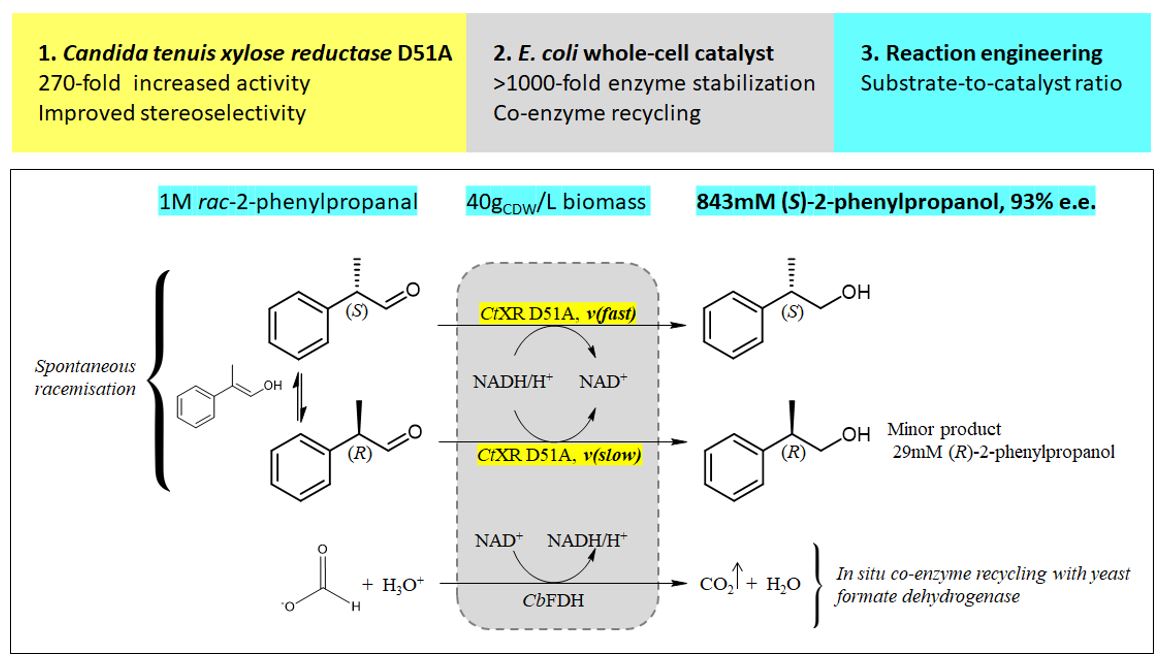
Reduction of rac-2-phenylpropanal by E. coli co-expressing CtXR D51A and a yeast formate dehydrogenase
Bioreductions were accomplished by lyophilized and rehydrated biomass or cell-free supernatant of engineered E. coli. The highly hydrophobic substrate formed a second phase in the reaction buffer above its solubility limit. Stirring led to droplet formation that were stabilized against coalescence in the presence of lyophilized biomass or supernatant.
100 mM rac-2-phenylpropanal
Conversions and product e.e. values for the reduction of 100 mM 2-phenylpropanal are summarized in Fig. 1. The amount of catalyst was varied between 4 and 40 gCDW/L. Comparison of lyophilized cells to cell-free supernatant used as catalyst was facilitated by using an equal amount of biomass for catalyst preparation in both cases. It has been previously shown that > 40 % of the total biomass activity is found as extracellular enzymes and hence in the supernatant (14). Conversions were similar for biomass and supernatant reflecting the severely impaired cell integrity caused by biomass lyophilization. Only at a low biocatalyst concentration of 4 gCDW/L, the supernatant led to significantly lower conversion (30 % lower). Supernatant and whole biomass showed equally high e.e. values of 95.3 % at 4 gCDW/L. Product enantiopurity and conversions increased gradually with increasing biocatalyst load. Notably, the e.e. values obtained with the supernatant decreased more strongly than the e.e. values from reductions with the whole biomass. At a catalyst load equal to 40 gCDW/L the e.e. values were 27 and 46 % for supernatant and whole biomass, respectively.
Optimization of 2-phenylpropanal bioreduction
Substrate concentration. Product concentration is the most important metric for bioprocesses to become compatible to chemical processes. Therefore, we increased the substrate concentration to 1 M at varying catalyst concentrations (lyophilized, rehydrated biomass) (Fig. 2, Table 3). The effects of increased catalyst loading on conversion and e.e.-values showed a similar but less pronounced trend compared to bioreductions with 100 mM substrate. A catalyst loading of 20 gCDW/L led to an e.e. value of 95 % compromised by a conversion of 28 %. Increase of the catalyst loading to 40 gCDW/L decreased the e.e. value to 93 % at 56 % conversion. A substrate concentration of 2 M turned out as too high, only low conversions of ~ 15 % were reached (Table 3).
NAD + concentration. We added higher concentrations of the co-enzyme NAD+ to further push the reaction towards full conversion. At 12 and 14 mM of NAD+, conversions up to 99% were reached, again at e.e. values of 92–93 % (Fig. 2, Table 3).
Cyclodextrin addition. The addition of 75 mM 2-hydroxypropyl-β-cyclodextrin has previously shown to boost bioreductions based on the used catalyst (14). Here, the addition of 38, 75 or 115 mM 2-hydroxypropyl-β-cyclodextrin had no significant effect on bioreductions of 1 and 2 M 2-phenylpropanal (Table 3).
Fed-batch. The step-wise addition of substrate at 0, 2 and 4 h to a total substrate concentration of 1 M led to a 10 % increase of conversion (Table 3).
Recovery, isolated yield, reproducibility and by-products
Leis et al. (20) previously suggested that hydrophobic substrates and products remain in the cell sludge of the used biocatalyst. Here, a high excess of ethyl acetate was required for product extraction prior to analysis. Obtained product concentrations in bioreductions of 1 and 2 M were between 27 and 84 % as shown in Table 3.
Recovery. Substrate/product loss in the biomass was found to be < 15% under the conditions used.
Reproducibility. Reaction replicates (N = 7) of bioreductions with 40 gCDW/L and 6 mM NAD+ showed high reproducibility with a mean value of 62 % product and a standard deviation of 4 %. The enantiomeric excess was 93.3 ± 1.1 % e.e. The formation of broad peaks prevented quantification of the aldehyde substrate by chiral, reversed-phase HPLC. We therefore analyzed bioreduction samples additionally by chiral GC-FID.
Table 3
Conversions and product e.e.-values of rac-2-phenylpropanal reduction by lyophilized whole-cell catalyst. Effects of catalyst loading, substrate concentration, co-enzyme concentration and HBC-addition.a
|
Catalyst loading
(gCDW/L)
|
rac-2-Phenylpropanal
(M)
|
NAD+
(mM)
|
HBC
(mM)
|
(S)-Phenylpropanol (mM)
|
e.e. (S)-Phenylpropanol
(%)
|
|
20
|
1
|
3
|
0
|
276
|
95.1
|
|
20
|
1
|
3
|
38
|
456
|
94.1
|
|
20
|
1
|
3
|
75
|
494
|
93.3
|
|
30
|
1
|
3
|
0
|
425
|
95.0
|
|
30
|
1
|
3
|
38
|
496
|
94.2
|
|
30
|
1
|
3
|
75
|
455
|
94.2
|
|
40
|
1
|
3
|
0
|
564
|
93.4
|
|
40
|
1
|
6
|
0
|
619
|
93.3
|
|
40
|
1 M fed-batch
|
6
|
0
|
679
|
94.0
|
|
40
|
1
|
8
|
0
|
732
|
94.1
|
|
40
|
1
|
10
|
0
|
843
|
93.1
|
|
40
|
1
|
12
|
0
|
839
|
92.9
|
|
40
|
1
|
14
|
0
|
765
|
92.3
|
|
40
|
1
|
6
|
38
|
634
|
94.3
|
|
40
|
1
|
6
|
75
|
598
|
92.9
|
|
40
|
1
|
6
|
115
|
592
|
92.5
|
|
40
|
2
|
6
|
0
|
279
|
95.4
|
|
40
|
2
|
6
|
38
|
306
|
93.4
|
|
40
|
2
|
6
|
75
|
306
|
92.2
|
| aReaction time 48 h. |
By-products. The high reactivity of the substrate 2-phenylpropanal prompted investigation of possible by-products from chemical or bio-chemical reactions. It has been previously shown that acetophenone forms by oxygen-catalyzed degradation of rac-2-phenylpropanal (21). We found 7% of acetophenone to be formed maximally and only trace amounts of its enzymatic reduction product in bioconversions of 1 M phenylpropanal, using 40 gCDW/L cells and 6 mM NAD+ (22). The substrate is also in a chemical equilibrium between rac-2-phenylpropanal and its corresponding hydrates. The previously reported enzymatic oxidation of 2-phenylpropanal hydrates to the corresponding carboxylic acids was not observed (12, 23). No substrate-related enol or aldol was found in detectable amounts (for data of NMR analyses see the Supplementary data).
Isolated yield. To confirm the identity of the obtained product, hydrophobic compounds were extracted from two reaction mixtures containing 1 M 2-phenylpropanol (reaction volume 2 mL, 40 gCDW/L catalyst, 6 mM NAD+). An analytical yield of 78 % was determined by HPLC (product concentration). The solvent and unreacted 2-phenylpropanal (bp 92 – 94°) were removed under reduced pressure. Product composition was 86 % 2-phenylpropanol, 7 % acetophenone and 7 % ethyl acetate (extractant) by 1H-NMR. No 2-phenylpropanal was found. An isolated yield of 64 % was obtained (203 mg with a product content of 86 %). (For data of HPLC, GC and NMR analyses see the Supplementary data).
Literature survey
Reported enzymatic reductive kinetic resolution of rac-2-phenylpropanal studies are summarized in Table 4. Bioreductions of rac-2-phenylpropanal have been accomplished with free-floating enzymes (Table 4, entries 1–5, 8–10,12) and immobilized enzymes (entries 6,7,11). Previous studies aimed at probing bioreduction catalysts (free and immobilized oxidoreductases) in the kinetic resolution of rac-2-phenylpropanal (entries 4–11). Most enantioselective enzymes preferred the (S)-aldehyde (entries 1–8, 10). Rocha-Martín et al. (24) reported on an anti-prelog specific ADH from Thermus thermophilus HB27 (entry 11). Dong et al. (8) accomplished the evolution of ADHs for the formation of (S)- and (R)-alcohols by directed evolution of an ADH from Thermoanaerobacter brockii that displayed moderate prelog-type selectivity (entries 8,9). The used enzymes had to display not only high enantioselectivities but also sufficient stabilities in the presence of the substrate that can form adducts with groups on the enzymes (25). HLADH was used in most studies as it turned out to be enantioselective, stable in the presence of substrate up to a concentration of 165 mM, and useful in coupled substrate strategies (oxidation of cheap alcohols for NADH-recycling). All other examples of selective ADHs stem from thermophilic organisms and display intrinsically high stabilities towards adverse effects of the reaction media. Remarkably, the often-used host E. coli shows native activity towards 2-phenylpropanal (entry 13). Buffered solutions containing water-soluble co-solvents (also used as sacrificial substrate for NADH-recycling) were used frequently. The aqueous phase was required for the racemisation of the substrate. Grunwald et al. (11) tested HLADH in organic solvent and used isopropylether with 0.5 % buffer as reaction medium. A product concentration of 46 mM with 95 % e.e. was obtained, however at a conversion of 15 % (entry 7). Others used bi-phasic solvents (entries 4 and 6). The highest published product concentration of 82 mM was achieved in a buffer/isopropylether mixture (47:63) (entry 4).
Table 4
Literature survey of enzymatic reductive kinetic resolution of rac-2-phenylpropanal
|
Entry
|
Bioreduction catalyst,
NAD(P)H-recycling strategy
|
rac-2-phenylpropanal
|
Medium,
(auxiliary substrate)
|
Product
(Conversion)
|
Enantiopurity
|
Aim of the study
|
Ref.
|
| |
Horse liver ADH (Zn-containing ADH)
|
|
|
|
|
|
|
|
1
|
Free enzyme,
coupled substrate 1,4-butanediol
|
5 mM
|
Buffer pH 7.5, 1 % v/v CH3CN,
(2.5 mM 1,4-butanediol)
|
5 mM
(99 %)
|
95 % e.e. S
|
Probing the enzyme's co-enzyme recycling ability using 1,4-butanediol oxidation to lactone
|
26
|
|
2
|
Free enzyme,
coupled substrate ethanol
|
0.5 mM
|
Buffer pH 7.5,
(0.5 M ethanol)
|
0.38 mM
(75 %)
|
98 % e.e. S
|
Investigation of DKR including substrate racemization velocity
|
27, 10
|
|
3
|
Free enzyme,
phenylpropanal oxidation for NADH-recycling
|
75 mM
|
Buffer pH 7.5; 4 % v/v MTBE
|
23 mM
(31 %)
|
97 % e.e. S
|
Investigation of the biocatalytic asymmetric disproportionation (biocatalytic Cannizzaro reaction)
|
12, 28
|
|
4
|
Free enzyme,
coupled substrate ethanol
|
165 mM
|
Buffer, 63 % v/v isopropylether,
(0.6 M ethanol)
|
82 mM
(50 %)
|
96 % e.e. S
|
Probing the enzyme's enantioselectivity and co-enzyme recycling ability
|
29
|
|
5
|
Free enzyme,
coupled substrate ethanol
|
30 mM
|
Buffer pH 8,
(5% v/v ethanol)
|
28 mM
(93 %)
|
93 % e.e. S
|
One-pot, two-step reaction: rac-2-phenyl-1-propanol oxidation to rac-2-phenylpropanal followed by enantioselective bioreduction under dynamic conditions.
|
1
|
|
6
|
Immobilized enzyme,
coupled substrate ethanol
|
5 mM
|
Buffer pH 7.5, 50 % v/v hexane,
(0.5 M ethanol)
|
4.2 mM
(84 %)
|
> 98 % e.e. S
|
Characterization of the immobilized catalyst
|
30
|
|
7
|
Immobilized enzyme,
coupled substrate ethanol
|
300 mM
|
Isopropyl ether (saturated with buffer), 0.5 % buffer pH 7.0,
(1 M ethanol)
|
46 mM
(15 %)
|
95 % e.e. S
|
Probing the enzyme's substrate scope and enantioselectivity in organic solvent
|
11
|
| |
Thermostable ADHs (enzyme superfamily)
|
|
|
|
|
|
|
|
8
|
Free Thermoanaerobacter brockii LG296 ADH mutant (Zn-containing ADH),
coupled substrate isopropanol
|
30 mM
|
Buffer pH 7.4,
(20 % v/v isopropanol)
|
23 mM
(75 %)
|
95 % e.e. S
|
Development of enantioselective mutants
|
8
|
|
9
|
Free Thermoanaerobacter brockii LG277 ADH mutant (Zn-containing ADH),
coupled substrate isopropanol
|
10 mM
|
Buffer pH 7.4,
(20 % v/v isopropanol)
|
7.5 mM
(75 %)
|
92 % e.e. R
|
Development of enantioselective mutants
|
8
|
|
10
|
Free Sulfolobus solfataricus ADH-10 (Zn-containing ADH), coupled substrate ethanol
|
5 mM
|
Buffer pH 9,
(5 % ethanol)
|
3.7 mM
(74 %)
|
98 % e.e. S
|
Probing the enzyme's substrate scope and enantioselectivity
|
9
|
|
11
|
Immobilized Thermus thermophilus ADH (short-chain dehydrogenases/reductase),
coupled enzyme yeast formate dehydrogenase
|
1 mM
|
Buffer pH 7, 5 % v/v CH3CN
(0.1 M formic acid)
|
1 mM
(100 %)
|
71 % e.e. R
|
Characterization of the immobilized catalyst
|
24
|
| |
Origin of enzyme not stated
|
|
|
|
|
|
|
|
12
|
Free Evo-1.1.200 from Evocatal
coupled substrate ethanol
|
30 mM
|
Buffer pH 9,
(5% v/v isopropanol)
|
29 mM
(95 %)
|
89 % e.e. R
|
One-pot, two-step reaction: rac-2-phenyl-1-propanol oxidation to rac-2-phenylpropanal followed by enantioselective bioreduction under dynamic conditions.
|
1
|
| |
Whole-cell catalyst
|
|
|
|
|
|
|
|
13
|
E. coli JM109, NAD(P)H-recycling by native microbial metabolism
|
22 mM
|
M9 medium, 30 % v/v organic phase (9:1 isopropylether:isooctane)
|
4.5 mM
(48 %)
|
~ 50 % e.e. (? unknown)
|
Probing the host background activity
|
31
|
|
14
|
CtXR D51A mutant (aldo-keto reductase)
coupled enzyme yeast formate dehydrogenase
|
1000 mM
|
Buffer pH 7.5,
(1.05 M formic acid)
|
843 mM
(98.8 %, but product and substrate loss)
|
93.1 % e.e. S
|
Process optimization regarding enantiopurity and yield
|
This work
|
{kind=link}
{kind=link}