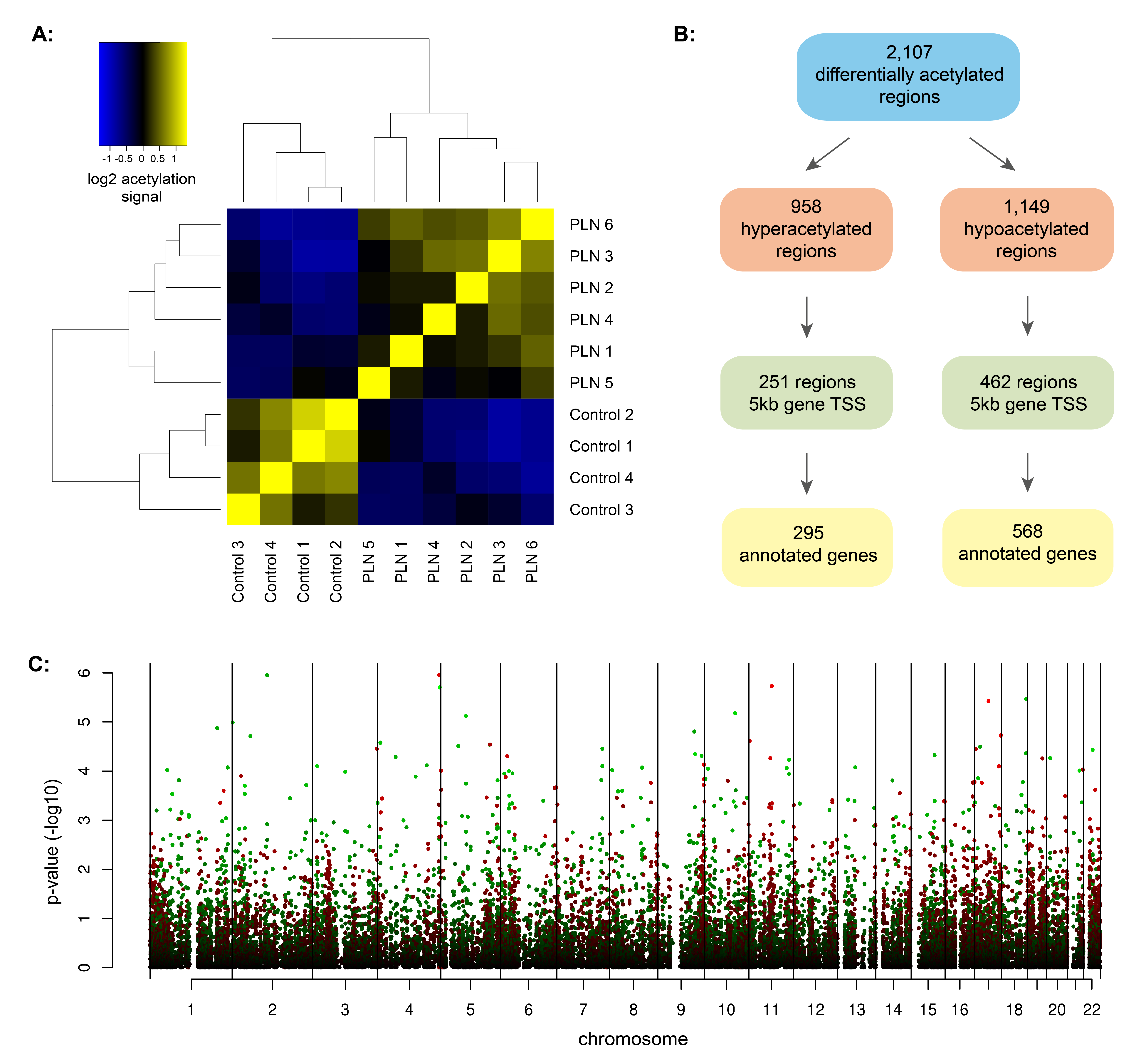
Identification of histone acetylation changes in PLN-R14del cardiac tissue
H3K27ac histone acetylation regulates gene transcription and contributes to phenotypic responses in heart diseases.25 Therefore, we performed H3K27ac ChIP-seq to study histone acetylation changes in 6 PLN-R14del versus 4 control hearts (Fig. 1A and Table S1A). We identified 28,149 ± 9,538, and 25,721 ± 8,460 H3K27ac enriched regions within PLN-R14del and control hearts, respectively. We subsequently combined regions that were identified in at least two independent samples into a set of 23,356 regions to assess differentially acetylated regions between control and PLN-R14del groups. In total, 2,107 autosomal regions showed differential H3K27ac levels between PLN-R14del and control hearts (Fig. 1B and Table S2A). Compared to controls, regions with higher H3K27ac levels in PLN-R14del hearts are referred to as hyperacetylated regions (n = 1,149) and regions with lower H3K27ac levels in PLN-R14del hearts are referred to as hypoacetylated regions (n = 958, Fig. 1C and Fig.S1).
Genes annotated to differentially acetylated regions in PLN-R14del cardiac tissues are enriched in metabolic pathways
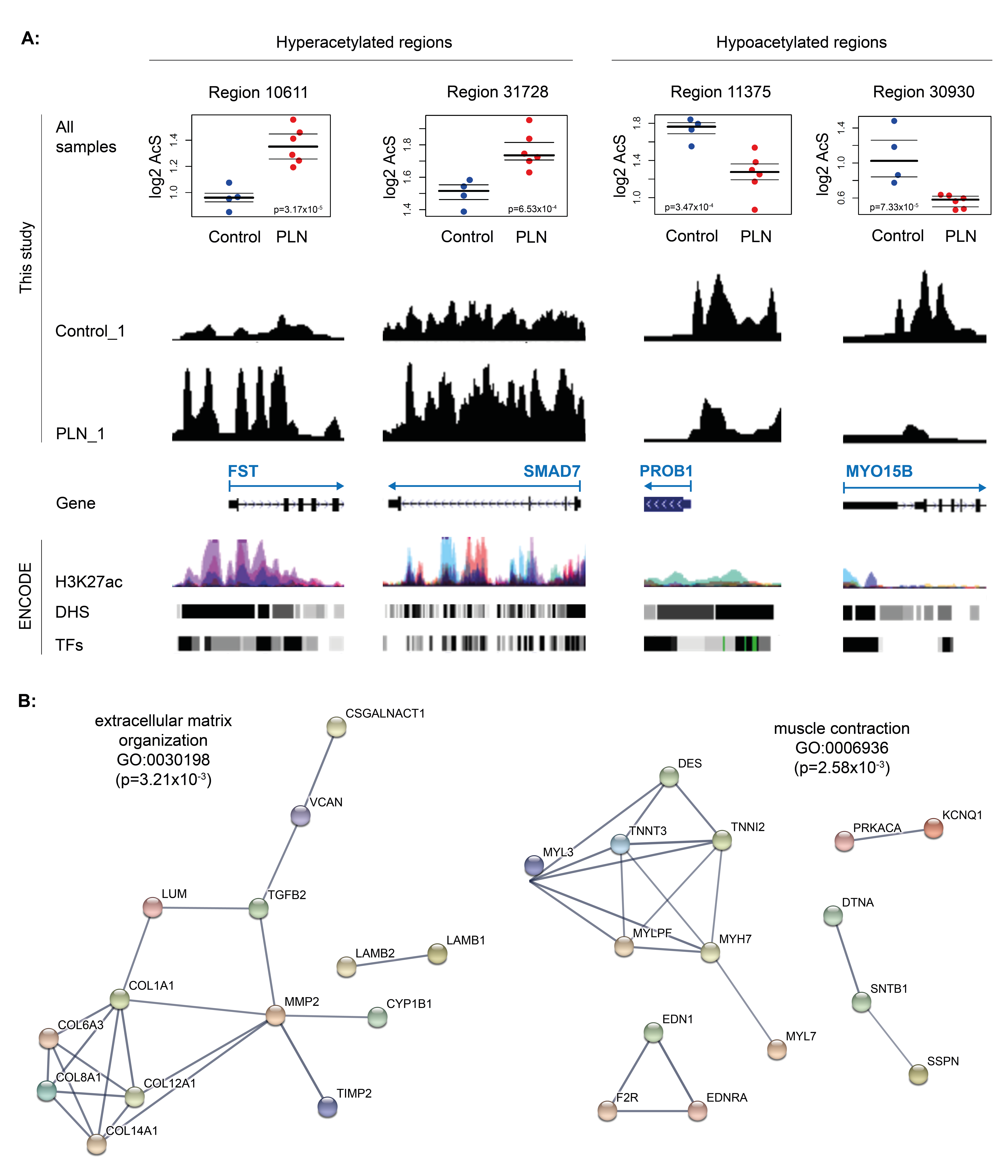
To identify genes potentially regulated by the differentially acetylated regions, we focused on regions in close vicinity to promoters and annotated genes 5,000 bases up- and downstream from the transcription start site of a gene as used previously (Fig. 1B).25,26 Out of 968 hyperacetylated regions, 295 genes were identified in close vicinity to 251 hyperacetylated regions, and out of 1,149 hypoacetylated regions, 568 genes were identified in the close vicinity to 462 hypoacetylated regions (Fig. 1C, Table S2B and 2C). To examine which biological processes and pathways are affected, we performed gene set enrichment analysis using genes annotated to differentially acetylated regions. We observed that hyperacetylation-related genes were mostly involved in fibrosis, (cardiovascular) development, and chromatin assembly (Table S2D and Fig.S2), while hypoacetylation-related genes were related to metabolism (Fig. 1D and Table S2E).
The transcription factor binding motifs (TFBMs) overrepresented in hypoacetylated regions are enriched in metabolic pathways
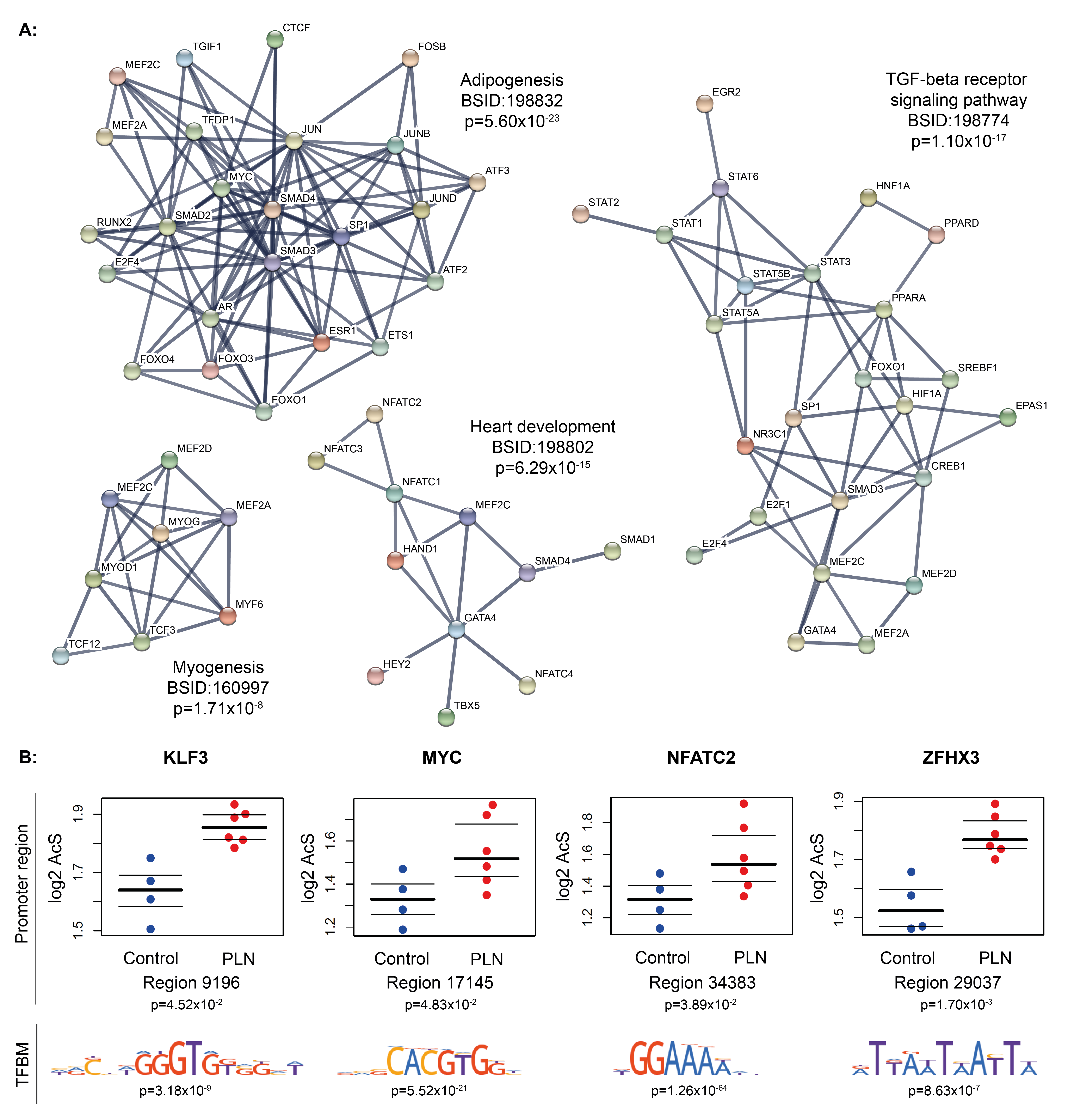
To identify possible upstream acting transcription factors (TFs), which regulate genes involved in the pathogenesis of the disease, we studied the overrepresentation of TFBMs in differentially acetylated regions. By using the DNA sequences of all differentially acetylated regions in PLN-R14del versus control hearts (Fig. 1B), we detected enrichment in 202 TFBMs and annotated them to 200 TF-encoding genes (Table S3A). Consistently, several of the most enriched biological processes annotated to TFs pointed towards altered metabolism, such as adipogenesis and mitochondrial structure (Table S3B and Fig.S3). Notably, PPARA, a major regulator of cardiomyocyte lipid metabolism, particularly FAO, was also annotated from enriched motifs together with other interacting TFs (Fig. 1E). Therefore, we further investigated the localization of PPARA in cardiac tissues by immunofluorescence staining and observed a significant decrease in the nuclear PPARA signal of PLN-R14del cardiomyocytes versus the controls, whereas the PPARA signal in non-myocyte cells remained comparable between PLN-R14del and control hearts (Fig. 1F).
Hypoacetylated regions associated with metabolic pathways specific for PLN-R14del cardiomyopathy as compared to other cardiomyopathies
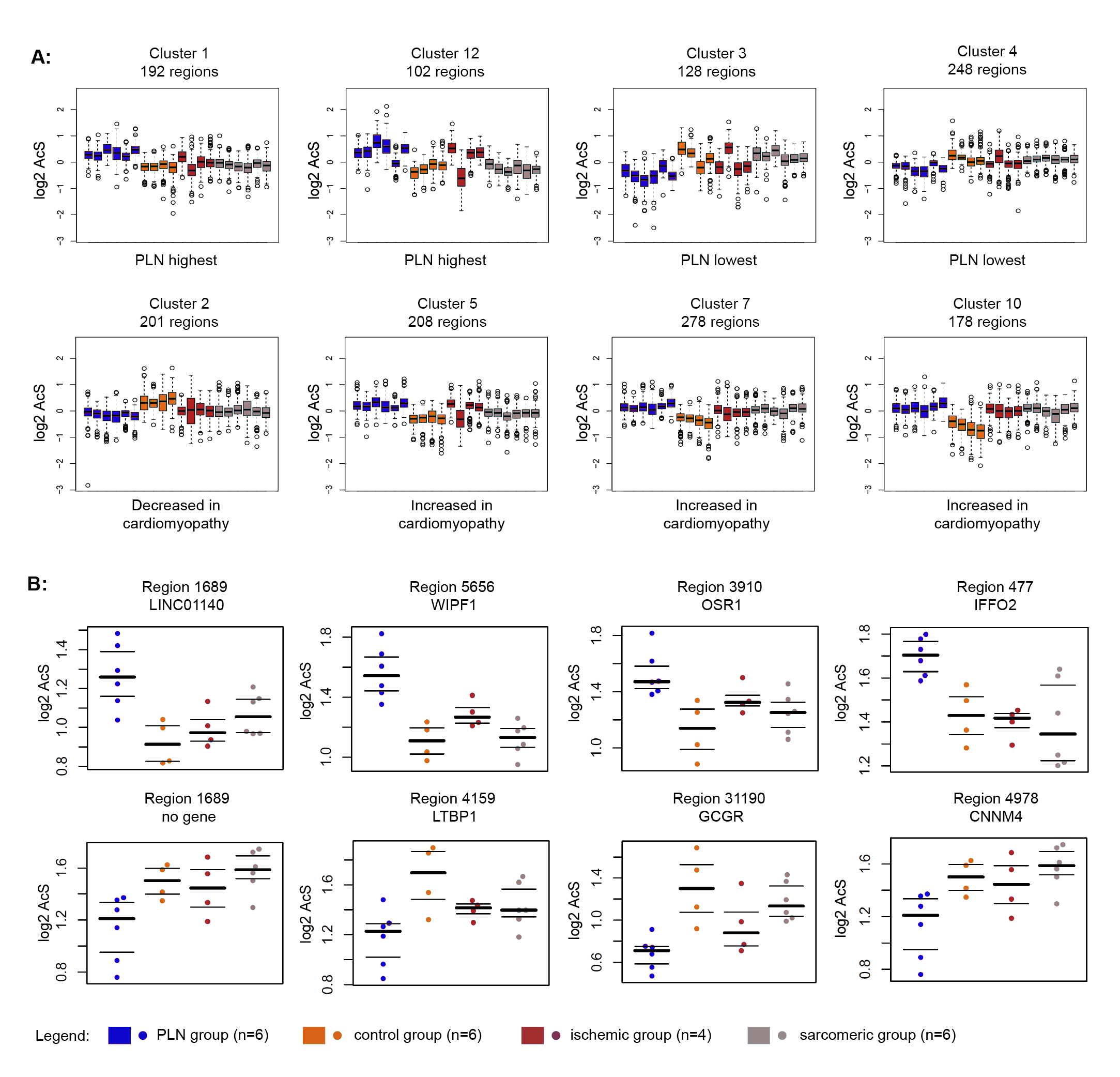
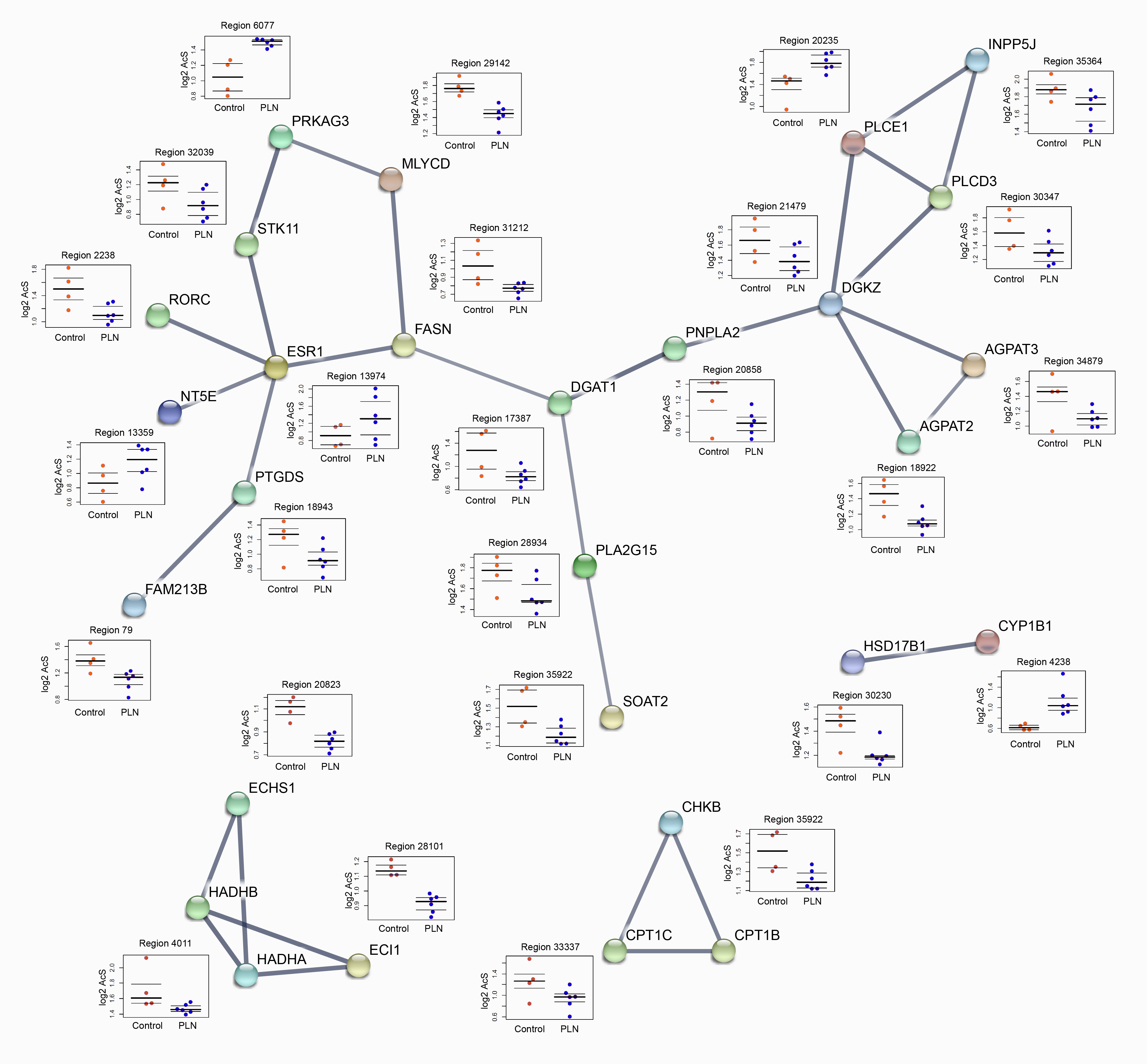
Besides non-failing control hearts, we also compared PLN-R14del hearts with other cardiomyopathies, including ischemic cardiomyopathy (n = 4) and non-ischemic dilated cardiomyopathy (sarcomeric group, n = 6, Fig. 1G). K-mean clustering analysis revealed four PLN-R14del-specific clusters when compared to other cardiomyopathy groups and the controls (Fig. 1H, Fig.S4, and Table S4A,). Genes located in the vicinity of these PLN-R14del-specific clusters were again highly enriched in metabolic signalling (Table S4B and 4C, and Fig.S5). Examples of metabolic genes, including HADHA/HADHB, SLC25A20, PDK2, and CPT1B, which were annotated in PLN-R14del-specific hypoacetylated clusters were shown in Fig. 1I. Combined, we detected differentially acetylated regions that distinguish PLN-R14del from other types of cardiomyopathies and they annotated metabolic-related genes were profoundly affected.
Enriched metabolic pathways by differentially expressed genes in PLN-R14del hearts
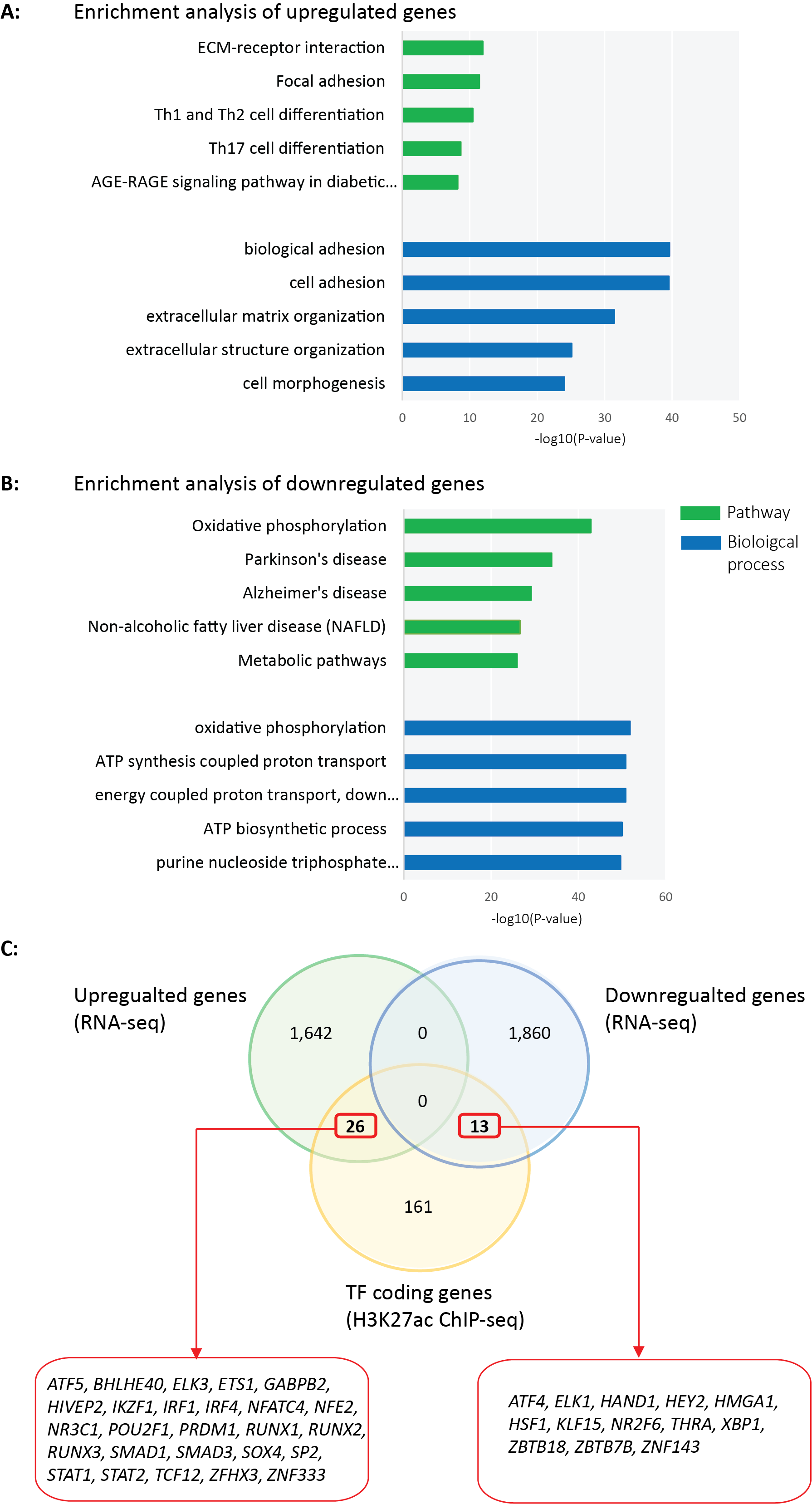
Besides differentially acetylated regions, we obtained 1,668 up- and 1,873 downregulated genes in PLN-R14del versus control hearts using RNA-seq (Fig. 2A and Fig. 2B, Table S5A). In line with the well-established suppression of SERCA2A/ATP2A2 at the protein level,6 we showed its suppression at the mRNA level for the first time. Additionally, metabolic genes, such as HADHA and HAHDB, which were annotated from PLN-R14del-specific hypoacetylated clusters, also showed significantly lower mRNA levels in PLN-R14del versus control hearts (Fig. 2C). We further demonstrated decreased HADHA and SERCA2A/ATP2A2 protein levels in PLN-R14del versus the control heart by immunofluorescence staining (Fig. 2D). Consistent with enriched biological processes and pathways by annotated genes from differentially acetylated regions, fibrosis, (cardiovascular) development, and chromatin assembly were enriched by upregulated genes (Table S5B) and metabolism (oxidative phosphorylation, ATP metabolic process, metabolic pathways, etc.) were enriched by downregulated genes (Fig. 2E and Table S5C).
Notably, among 200 TFs annotated from differentially acetylated regions, 39 TFs showed significantly altered mRNA levels in PLN-R14del versus control hearts, including 26 up- and 13 down-regulated TF-coding genes. Enriched protein-protein-interaction network by 39 TF-coding genes again suggests (negatively) affected metabolism (Fig. 2F). Thus, we identified a panel of upstream TFs and downstream targets in metabolic processes, which were disrupted in PLN-R14del cardiomyopathy.
Differentially expressed genes in PLN-R14del hiPSC-CMs suggest altered lipid metabolic pathways
Monolayers of hiPSC-CMs have been used to study the molecular mechanisms underlying several major cardiomyopathies, including ion-related, structural, and metabolic cardiomyopathies. However, the physiological immaturity of hiPSC-CMs severely limits their utility as a prediction model for adolescent genetic myopathies. To improve the cardiac immaturity limitation, we cultured hiPSC-CMs for 160 days in a maturation media designed to provide oxidative substrates adapted to the metabolic needs27 (Table S6). These long-term cultured hiPSC-CMs showed well-developed structural and mitochondrial organization as stained by the sarcomeric and mitochondrial marker (Fig.S7A). Hereafter, we studied the transcriptome profiles in PLN-R14del and healthy control hiPSC-CMs (Fig. 3A and Table S1B).
First, we examined markers of cytoskeletal components (ACTN1, TNNT2, MYH7, and MYL2), ion channels (KCNA5 and KCNJ4), and mitochondrial components (ATP5F1A and HSP60), and lipid metabolism (PPARA, ACAT1, FABP3, and Nile red staining). In general, we observed an increase in mRNA levels of most markers from short- and long-term (25 and 160 days, respectively) cultured PLN-R14del and healthy control hiPSC-CMs and confirmed these observations at the protein level by immunofluorescence staining (Fig. 3B and Fig. 3C). Therefore, we extended the culture time of hiPSC-CMs in the following experiments, which showed an overall improved maturation status but a more distinguishable metabolism-phenotype between PLN-R14del and control hiPSC-CMs.
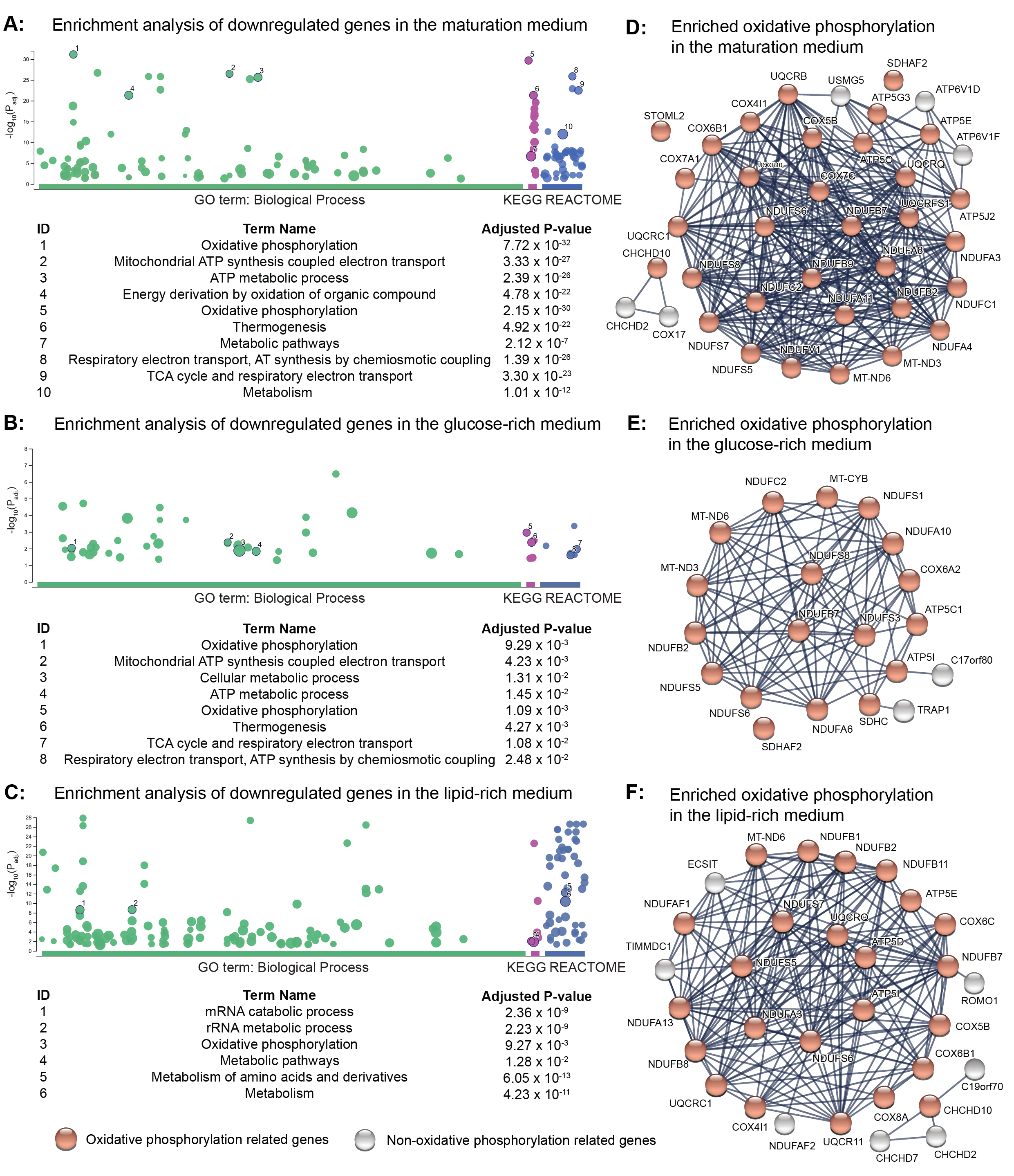
Since our data obtained from PLN-R14del hearts indicated a disrupted lipid metabolism, we cultured control and PLN-R14del hiPSC-CMs to further elucidate the metabolic activities in three culture media containing different amounts of glucose and lipids, which are the two main metabolic substrates for cardiomyocytes (Fig. 3A). In total, we identified 952, 1,321, and 2,104 differentially expressed genes in PLN-R14del versus control hiPSC-CMs cultured in the maturation, the glucose-rich, and the lipid-rich medium, respectively (Table S7A-C). Notably, regardless of culture media, pathway enrichment analyses using downregulated genes in PLN-R14del versus control hiPSC-CMs consistently pointed towards metabolic activities, such as oxidative phosphorylation/GO:0006119 (Fig. 3D, Fig.S8A-C, and Table S7D-F). However, it is important to note that the metabolic genes involved in the enriched biological processes (i.e., oxidative phosphorylation), which were shared among three conditions, were not the same (Fig.S8D-F). Similarly, regardless of culture media, pathway enrichment analyses using upregulated genes in PLN-R14del versus control hiPSC-CMs consistently pointed towards fibrosis and (cardiovascular) development, which were in line with the results obtained from the ex vivo human cardiac tissues (Table S7D-F).
Disturbed fatty acid oxidation (FAO) and metabolic flexibility in PLN-R14del hiPSC-CMs
Besides the downregulated transcriptional regulation of lipid metabolism in PLN-R14del cardiomyopathy, a significantly lower cellular metabolic activity/viability was also observed in PLN-R14del versus control hiPSC-CMs (Fig. 4A). This suppression remained when excessive glucose or FAs were given to the cells (Fig.S7B-C). To further elucidate whether the obtained transcriptomic data could predict affected metabolism in PLN-R14del cardiomyopathy, we compared the FAO metabolism, a key metabolic program in cardiomyocytes (Fig. 4B), by evaluating mitochondrial respiration via ETO-inhibited FAO and 2-DG-inhibited glycolysis using the Seahorse analysis (Fig. 4C).
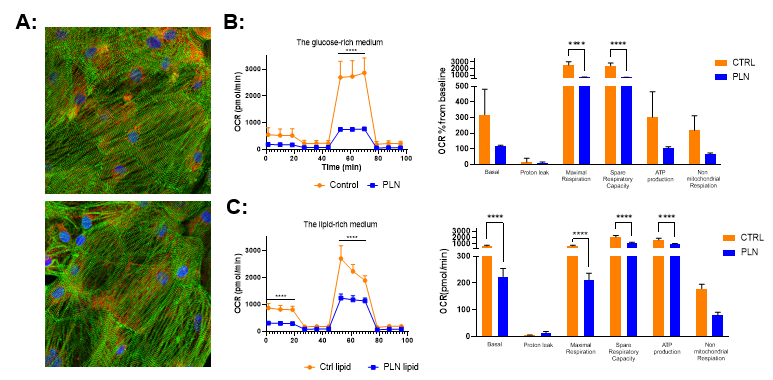
In the maturation medium, which contains both glucose and FAs, we observed a comparable oxygen consumption rate (OCR) between PLN-R14del and control hiPSC-CMs at the baseline level and after ETO-inhibited FAO (Fig. 4E), suggesting a similar FAO-dependence of both groups. However, after blocking glycolysis by 2-DG, we observed an increased OCR in control hiPSC-CMs, whereas the OCR of PLN-R14del hiPSC-CMs continued to decline significantly, suggesting control hiPSC-CMs are less dependent on glycolysis and have more metabolic adaptive characteristics than PLN-R14del hiPSC-CMs.
Similarly, in the glucose-rich medium, OCR was comparable between PLN-R14del and control hiPSC-CMs at the baseline level (Fig. 4F). After blocking glycolysis, a significantly higher OCR was observed in control versus PLN-R14del hiPSC-CMs, once again suggesting better metabolic flexibility and substrate utilisation in control hiPSC-CMs.
Notably, in the lipid-rich medium, the basal OCR was significantly lower in PLN-R14del versus control hiPSC-CMs (Fig. 4G), implying an impaired FAO in the PLN-R14del hiPSC-CMs at the beginning. Although OCRs of both PLN-R14del and control hiPSC-CMs decreased after FAO- and glycolysis-blockages, OCRs remained significantly lower in PLN-R14del hiPSC-CMs than in the controls. These findings suggest that despite the presence of excess lipids in the medium, PLN-R14del hiPSC-CMs are less capable of utilizing their FAO metabolism to produce the required energy, therefore being more glycolysis-dependent.
Higher glucose dependency in PLN-R14del hiPSC-CMs
Glucose metabolism, another major metabolic program in cardiomyocytes (Fig. 4B), was also studied. We measured the extracellular acidification rate (ECAR) to study the activity of the glycolytic pathway in PLN-R14del and wild-type hiPSC-CMs by manipulating the FAO and glucose metabolism using ETO and 2-DG respectively (Fig. 4D).
In the maturation medium, basal ECAR levels were comparable between both groups, but it became significantly lower in PLN-R14del versus control hiPSC-CMs after FAO- and glycolysis-blockages (Fig. 4H), indicating a higher glycolysis dependency of PLN-R14del hiPSC-CMs. A significantly higher glycolytic reserve was also observed in PLN-R14del hiPSC-CMs than in the controls.
In the glucose-rich medium, ECAR levels remained comparable between both groups at the basal level, after blocking FAO, and after blocking glycolysis (Fig. 4I). However, the decline of ECAR after glycolysis-blockage was more profound in PLN-R14del versus control hiPSC-CMs, suggesting a higher glycolysis-dependence and a higher glycolytic reserve of PLN-R14del hiPSC-CMs.
In the lipid-rich medium, basal ECAR levels were comparable between both groups (Fig. 4J). Interestingly, a higher ECAR was shown in PLN-R14del versus control hiPSC-CMs after FAO-blockage, which decreased profoundly after glycolysis-blockage, again, confirming the higher glycolysis-dependency in PLN-R14del hiPSC-CMs. These results imply our in vitro maturation-induced PLN-R14del hiPSC-CM mimics the heart failure-related metabolic alterations consisting of the energy production reduction by mitochondria through oxidative phosphorylation and an increase in (anaerobic) glycolysis.
Intracellular lipid droplet accumulation is a key feature of PLN-R14del cardiomyopathy
Data acquired from the cardiac tissues and/or hiPSC-CMs at DNA, RNA, protein, and functional levels consistently pointed towards altered metabolism in PLN-R14del versus control groups, particularly FAO. Since lipid accumulation is known as the hallmark of impaired FAO, we examined the lipid accumulation in cardiac tissues and hiPSC-CMs.
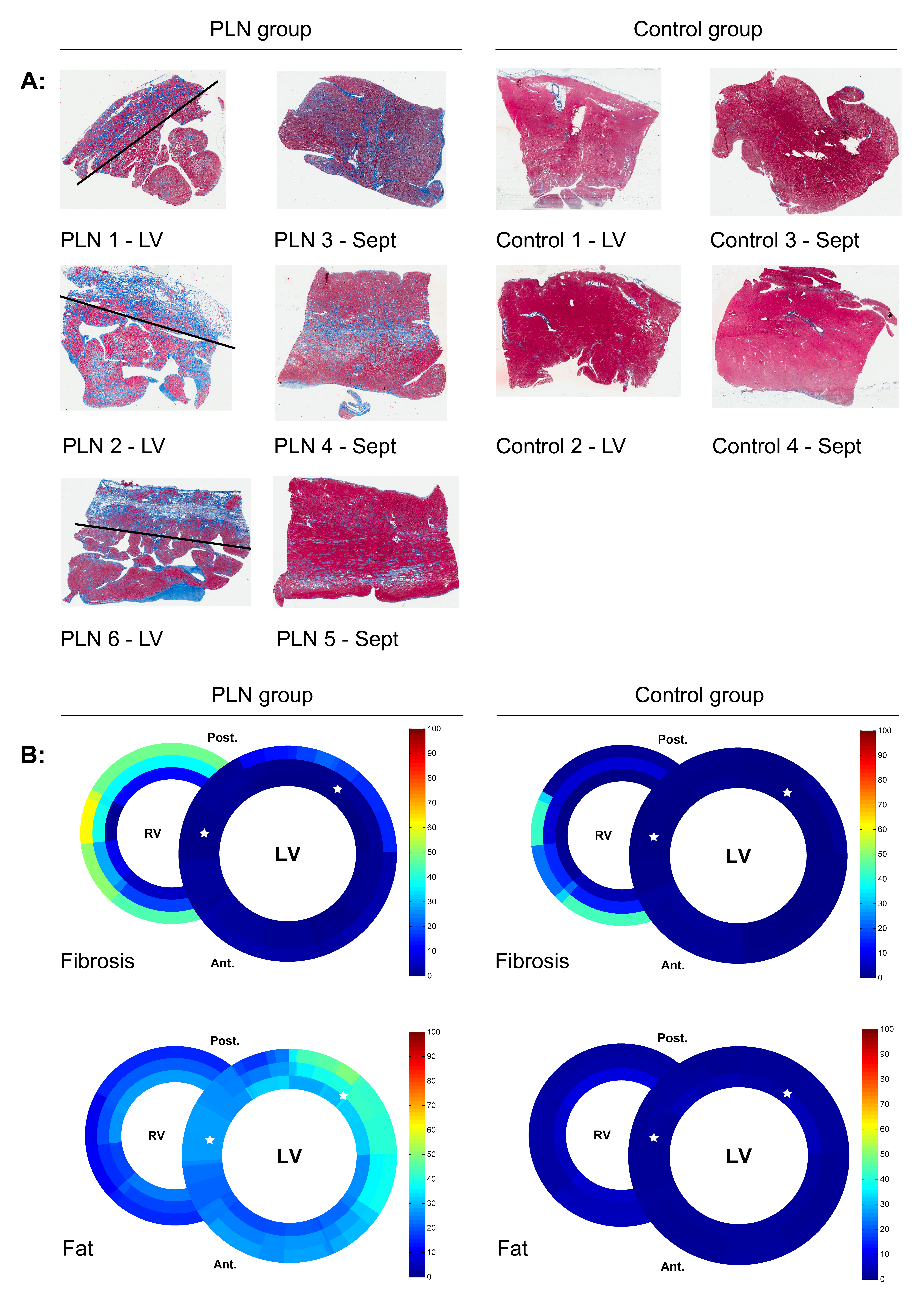
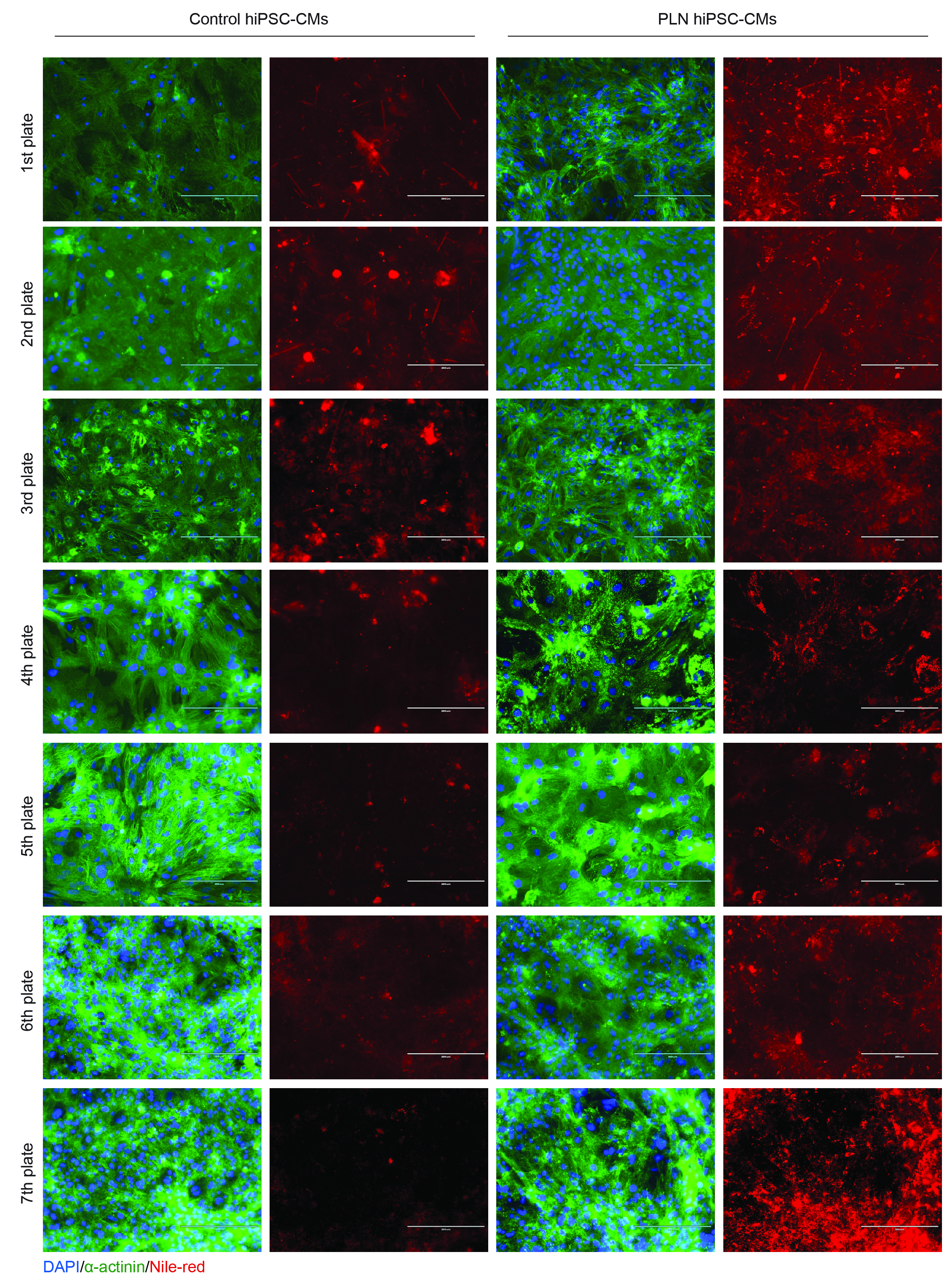
First, we performed a digital quantification in heart slices assessing the percentage of adipose tissue, which showed an increased adipocyte deposition in PLN-R14del versus control hearts (Fig.S8). Next, we used Nile red to localise intracellular lipid droplets in snap-frozen and paraffin-embedded tissues and observed a more frequent perinuclear accumulation of lipid droplets in PLN-R14del cardiac tissue than in the control (Fig. 5A). Transmission electron microscopy revealed a significantly lower mitochondrial length (aspect ratio) and a significantly higher accumulation of intracellular lipid droplets in PLN-R14del versus control hearts (Fig. 5B and Fig. 5C), suggesting impaired mitochondrial FAO. In line with the hearts, highly accumulated intracellular lipid droplets were observed in PLN-R14del versus control hiPSC-CMs, regardless of different culturing media (Fig. 5D). Combined, these findings suggested intracellular lipid droplet accumulation is a key feature in PLN-R14del cardiomyopathy.
CRISPR/Cas9-based correction of PLN-R14del attenuated intracellular lipid accumulation
We used CRISPR/Cas9-based gene editing to correct PLN-R14del mutation28 and observed a significant reduction of intracellular lipid droplets in R14del-corrected hiPSC-CMs (isogenic control, Fig. 6A). Interestingly, hiPSC-CMs derived from an asymptomatic PLN-R14del carrier and the homozygous hiPSC-CMs showed a comparable amount of the lipid droplets as in PLN-R14del hiPSC-CMs. These findings suggested a tight relationship between PLN-R14del mutation and impaired lipid metabolism, thereby leading to intracellular lipid droplet accumulation.
PPARA-targeted drug increased Ca2+ handling and mitochondrial trifunctional protein levels in PLN-R14del hiPSC-CMs
We also applied a PPARA agonist (bezafibrate) to further investigate PPARA-mediated FAO in PLN-R14del cardiomyocytes. First, we measured the Ca2+ transient in control and PLN-R14del hiPSC-CMs with and without bezafibrate treatment. We observed significantly increased rise and decay time in treated versus untreated PLN-R14del hiPSC-CMs (Fig. 6B), whereas the treatment did not affect Ca2+ handling in control hiPSC-CMs. Next, we examined HADHA and HADHB levels, encoding the mitochondrial trifunctional protein involved in the FAO pathway, which showed suppressed histone acetylation and transcriptional levels in PLN-R14del versus control hearts (Fig. 1I and Fig. 2C). We confirmed the suppression of HADHA and HADHB in long-term cultured untreated PLN-R14del cardiomyocytes and further showed elevated HADHA and HADHB levels in bezafibrate-treated PLN-R14del cardiomyocytes (Fig. 6C). Whereas, the treatment did not alter HADHA and HADHB levels in control hiPSC-CMs. Combined, these findings suggested disturbed HADHA/HADHB-involved FAO in PLN-R14del hiPSC-CMs, the potential association between PPARA-mediated FAO and Ca2+ handling, and the role of PPARA as a promising therapeutic target in PLN-R14del cardiomyopathy.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}