The metabolic syndrome (MetS), which includes type 2 diabetes mellitus, hypertension insulin resistance, dyslipidemia, non-alcoholic fatty liver disease (NAFLD), and the obstructive sleep apnea syndrome, are most likely to caused by obesity, especially central obesity [26]. The phytophenols are being studied as natural, potential therapeutic agents for a number of diseases and metabolic syndromes. Gallic acid (GA), which has curative effects on various metabolic syndromes, has been used in our study. According to our observations, animals on high fat diet (HFD) acquire more weight but consistently utilised less food as compare to control. Also the oral administration of GA in HFD-induced weight gain in female mice has brought about considerable effect on body weight without influencing food intake. Our observation supports the findings of Doan et al., who have confirmed that GA has a positive effect on weight and energy metabolism. Notably, GA activates the AMPK signalling pathway to regulate body weight in diet induced obesity [27].
The various physiological process like biosynthesis of various hormones in the body, energy homeostasis and energy expenditure is directly linked with lipid metabolism. Lipid serves as a substrate for numerous reactions, a signalling molecule, and a biomarker. Dyslipidemia and related comorbidities are brought on by dysregulation of the lipid metabolism homeostasis [28]. Triglyceride (TG) and cholesterol levels in this study are noticeably higher in the HFD group. The GA treatment in the HFD group improved TG and cholesterol levels over the course of 60 days, demonstrating hypolipidemic activity. Bak et al. [29] reported similar outcomes for improved triglyceride and cholesterol by gallic acid following HFD exposure for two weeks in C57BL/6 mice [29]. Increased TG production is one of the root causes of insulin resistance syndrome [30]. Importantly, increased fasting and postprandial TG, along with a low HDL-C and high proportion of small dense LDL, are the defining characteristics of dyslipidemia in obesity. As it will result in a slow clearing of the TG-rich lipoproteins, hypertriglyceridemia condition is generated in the body which could be the primary factor of some other lipid anomalies [31]. Moreover, the discrepancies in cholesterol metabolism may be due to underlying insulin effects which is linked to the distribution of fat as seen in obesity and indicates new aspects of the metabolic interaction among cholesterol and insulin [32]. Along with the glucose metabolism, increased cholesterol synthesis and low cholesterol uptake are associated with insulin resistance. A connection between cholesterol metabolism and insulin resistance may be provided by the regulatory action of hyperinsulinemia on cholesterol synthesis [33]. Insulin has been shown to activate liver X receptors (LXRs), which in turn activate genes that at least partially regulate cholesterol synthesis and lipogenesis [34]. This LXR-mediated combined regulation of cholesterol synthesis and lipogenesis through interactions of steroid regulatory elements proteins [35], therefore correlate insulin and cholesterol.
Notably, in our study, GA in the HFD group for 60 days led to noticeably lower serum glucose concentrations than the HFD alone group without any discernible change in daily dietary intake, supporting the research by Latha and Daisy showing the antidiabetic potential of GA against blood sugar. Our findings demonstrate that in the IPGTT, GA significantly controlled the glucose level by rapidly eliminating blood glucose in the HFD + GA group, indicating the anti-diabetic property of GA for controlling blood glucose level and preserving glucose homeostasis. Paraiso et al. have also shown improvement in metabolic profile along with glucose tolerance and insulin sensitivity with oral administration of GA [36]. Similarly, in our study the administration of gallic acid in the HFD + GA group significantly reduced insulin and HOMA IR in comparison to the elevated levels of the same in HFD female mice, indicating the ameliorative role of GA in HFD-induced insulin resistance.
The increased ectopic fat accumulation causes metabolic dysregulation, including adiposity, which is a significant risk factor for the emergence of insulin resistance [37]. The current study demonstrated a relationship between high fat diet and adipocyte expansion in VAT, which may be caused by deposition of lipid in adipocytes and is well marked by unusual weight gain and hyperlipidemia. Antidote GA greatly enhances lipid metabolism, which controls weight gain and fat storage. Our findings are supported by a study by Latha and Daisy that demonstrates the ability of GA to modify body composition, as well as its positive metabolic effects and antiobesity effects, including decreased visceral fat [38]. Additionally, the results of our study are in line with those of Bak et al., who found that GA reduced the hypertrophy of adipocytes in DOI mice.[29].
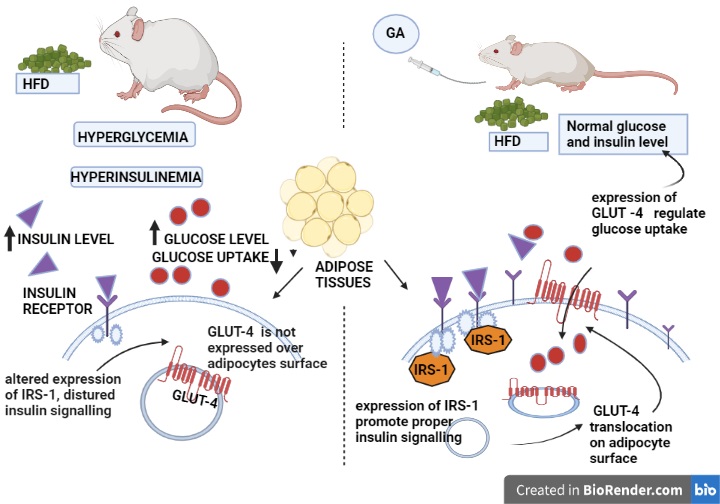
The current study demonstrates that visceral adipose tissue from the HFD and control group had significantly different expression levels of GLUT-4 mRNA. In this case, GLUT-4 regulation has changed as a impact of HFD on glucose metabolism. In visceral fat level of GLUT-4 mRNA expression in adipocytes may be a sign of a possible mechanism that plays a part in metabolic abnormalities. In consistency with our study numerous earlier studies have shown that obesity is linked to suppressed GLUT-4 expression, which results in an insulin resistance condition [39]. GLUT-4, which is specifically found on the cell membrane of adipose cells, is responsible for the majority of basal glucose transport. GLUT-4 predominantly regulates the transport of insulin and glucose through translocation and activation facilitated by the PI3K/phosphorylated protein kinase (p-Akt) pathway [40]. Transgenic mice which is over expressing or lacking GLUT4 have been shown to have reduced or enhanced entire body insulin sensitivity [41] highlighting its importance in glucose homeostasis. Interleukin-6 (IL-6) and tumour necrosis factor (TNF) and interleukin-6 (IL-6) are inflammatory cytokines produced by adipose tissue that have been linked to GLUT-4 expression reduction [42].
Adipose tissue undergoes a number of cellular processes that are influenced by insulin, such as cell division and proliferation, activation of amino acid and glucose uptake, suppression of lipolysis, remobilization of several membrane specific proteins, including insulin-like growth factor II receptor, and transferrin receptor and leptin synthesis [43, 44]. One of them, insulin-induced PI 3-kinase stimulation, is crucial in GLUT-4's movement from intracellular compartments to the surface of the cell [45]. In our research, we observed that HFD experimental females had low ISR-1 mRNA expression for 60 days. Our research is consistent with that of Anai et al., who found that rats fed with high-fat diet for specific duration, expressed significantly less (24%) IRS-1 in epididymal fat cells than did the control [11]. It has been demonstrated that IRS-1 along with IRS-2 takes part in the insulin signal transduction pathway, which induces GLUT4 translocation in adipocytes [46]. Moreover the administration of GA has improved the expression of GLUT-4 and IRS-1 in visceral adipocytes in HFD + GA. Our research confirms the findings of Gandi et al [47], who found that GA enhance the expression of GLUT-4 along with PI3K and pAKT over epididymal adipocytes in the of diabetic rats [47]. IRS-1 knockout mice exhibit lowered insulin-induced transport of glucose in insulin-responsive cells and tissues, such as adipose tissues and skeletal muscle, which is well supported by various experiments in rodent models and some in vitro studies that link alteration in expression of IRS1 to insulin resistance [48–50].
{kind=link}