Exposure to chronic stress led to significant changes in SIK1-CRTC1 signalling in PVN neurons
Since CSDS and CUMS are two well-validated and widely-used rodent models of depression (Antoniuk et al., 2019; Wang et al., 2021), they were applied together in this study. As shown in Figure 1A and 1B, exposure to CSDS and CUMS induced significant depression-like behaviours in C57BL/6J mice, as revealed by the FST, TST, sucrose preference test, and social interaction test. Meanwhile, exposure to CSDS and CUMS robustly enhanced the plasma levels of corticosterone and ACTH in mice (Figure 1A and 1B), representing a hyperactive HPA axis in response to chronic stress. Subsequently, western blotting was performed to detect the expression of SIK1-SIK3 in the total protein homogenates of PVN neurons. It was found that CSDS exposure downregulated SIK1 expression in the PVN by 55.4% ± 7.2%, whereas it had no influence on SIK2 or SIK3 in the PVN (Figure 1C). Similarly, CUMS exposure decreased SIK1 expression in the PVN by 58.9% ± 5.4% but induced no effects on SIK2 or SIK3 in the PVN (Figure 1D). The effects of chronic stress on the protein expression of CRTC1-CRTC3 in the PVN were also evaluated. As shown in Figure 1E and 1F, both CSDS and CUMS fully enhanced the level of nuclear CRTC1 in the PVN, but the levels of nuclear CRTC2 or CRTC3 were not affected. In contrast, chronic stress notably decreased the level of cytoplasmic phosphorylated CRTC1 (pCRTC1) in the PVN, while that of pCRTC2 was unchanged (Figure 1E and 1F). Interestingly, the total protein level of CRTC1 in the PVN in stressed mice was also significantly higher than that of control mice, whereas the levels of total CRTC2 and total CRTC3 did not increase (Figure 1E and 1F). As such, we focused on SIK1 and CRTC1 in the following studies.
Then, qRT‒PCR was performed to assay the mRNA levels of SIK1 and CRTC1 in PVN neurons. Figure S1A shows that CSDS and CUMS downregulated the SIK1 mRNA in the PVN by 55.8% ± 4.7% and 62.4% ± 8.6%, respectively. Figure S1A also reveals that CSDS and CUMS upregulated the CRTC1 mRNA in the PVN by 140.9% ± 9.5% and 166.3% ± 12.3%, respectively. Next, we examined the binding level of nuclear CRTC1 and CREB by Co-IP. It was found that chronic stress strongly promoted binding between nuclear CRTC1 and CREB in the PVN (Figure S2A and S2B), in accordance with the western blotting results of nuclear CRTC1 in Figure 1E and 1F. Moreover, we examined the neuronal distribution of CRTC1 in the PVN by immunofluorescence, with NeuN and DAPI as the nuclear markers. As shown in Figure 3A and 3B, compared with the control group, both CSDS and CUMS significantly increased CRTC1 single staining, CRTC1/NeuN double staining and CRTC1/NeuN/DAPI triple staining in PVN neurons, representing notable nuclear translocation and enhanced biosynthesis of CRTC1 under the effects of chronic stress. Taken together, these findings suggest that SIK1-CRTC1 signalling in PVN neurons is implicated in the pathophysiology of depression.
Both genetic knockdown of SIK1 and genetic overexpression of CRTC1 in the PVN induced a depression-like phenotype in naïve mice
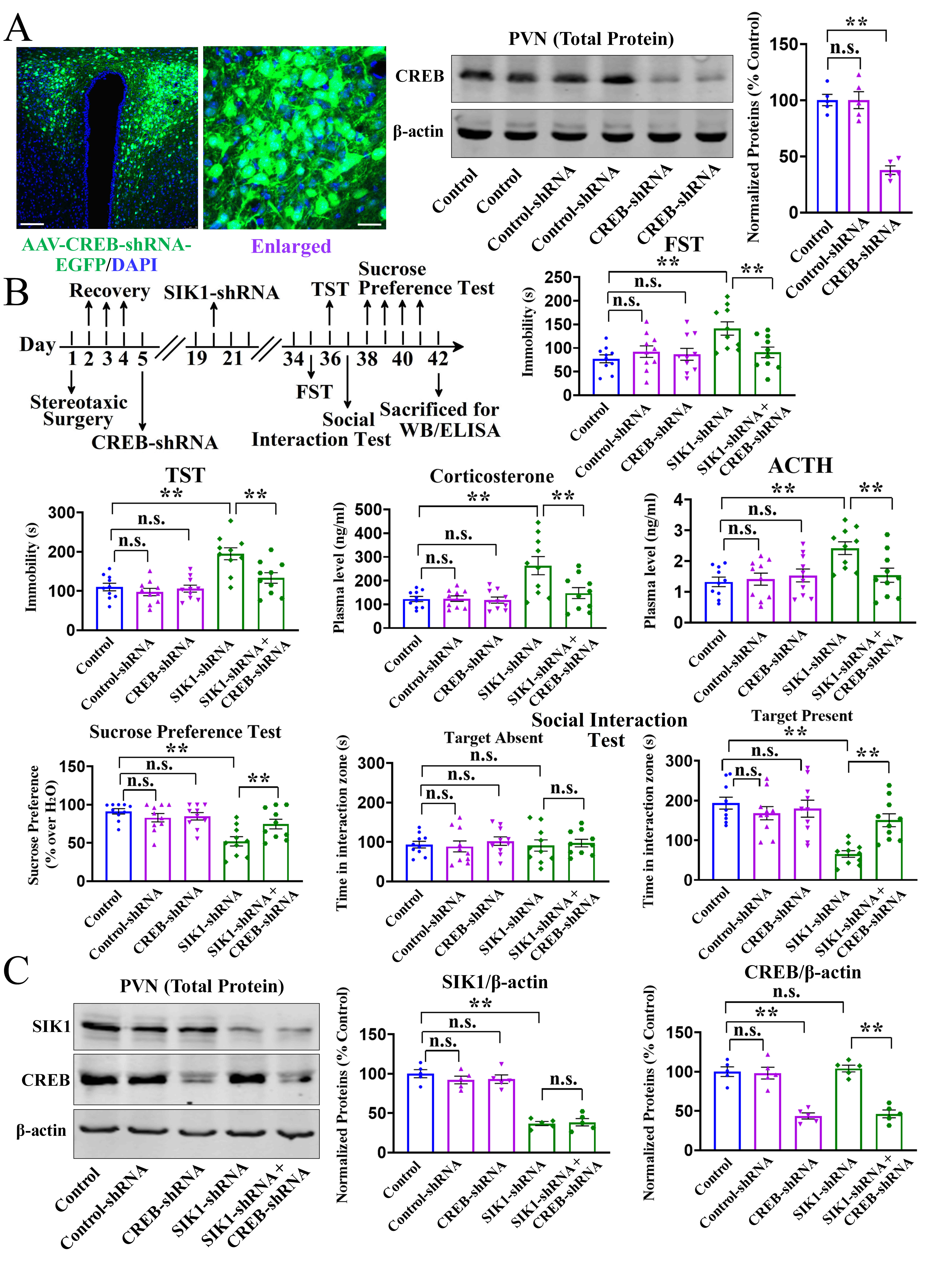
Since chronic stress downregulated SIK1 expression in the PVN, we then studied whether genetic knockdown of SIK1 in the PVN of naïve mice would simulate chronic stress, inducing depression-like behaviours. To achieve this purpose, an enhanced green fluorescent protein (EGFP)-containing AAV vector that expresses specific short hairpin RNAs (shRNAs) against SIK1 (AAV-SIK1-shRNA-EGFP) was generated. SIK1-shRNA or Control-shRNA was stereotactically and bilaterally infused into the PVN of naïve C57BL/6J mice, and after 14 d, numerous EGFP-positive neurons and notably decreased SIK1 expression were both observed in this region (Figure 2A). In behavioural tests, it was found that knockdown of SIK1 in the PVN largely enhanced the immobility of mice in both the FST and TST compared with that of the control group (Figure 2B), representing a behaviour of desperation and helplessness. To exclude the possible effects of SIK1 knockdown on mouse locomotor activity which may have led to a false-positive conclusion, the OFT was conducted. There was no significant difference in the number of squares the animals crossed in either the peripheral area or central area among all groups (Figure 2B). It was also found that mice with SIK1 knockdown in the PVN showed significantly less sucrose preference and social interaction than control mice (Figure 2B), representing behaviours of anhedonia and social avoidance. Meanwhile, the mice infused with SIK1-shRNA had remarkably higher levels of plasma corticosterone and ACTH than those of control mice (Figure 2B), representing a hyperactive HPA axis in response to SIK1 knockdown in the PVN. In contrast, the usage of Control-shRNA induced no effects on these evaluation indices.
Furthermore, knockdown of SIK1 in the PVN not only significantly increased the expression of nuclear CRTC1 (Figure 2C and 3C), total CRTC1 (Figure 2C and 3C), and CRTC1-CREB binding (Figure S2C) but also notably decreased the expression of cytoplasmic pCRTC1 (Figure 2C) in this region. More importantly, SIK1 knockdown remarkably enhanced the expression of CRH in the PVN (Figure 2C), consistent with the corticosterone and ACTH results in Figure 2B. Depression is accompanied by some critical pathological phenomena that can be attributed to the excessive release of glucocorticoids, such as decreased hippocampal neurogenesis and a downregulated brain-derived neurotrophic factor (BDNF) signalling cascade in the hippocampus and mPFC (Jiang et al., 2021; Liu et al., 2020; Masi and Brovedani, 2011; Mahar et al., 2014; Snyder et al., 2011). Interestingly, Figure 2D shows that the mice infused with SIK1-shRNA had significantly lower protein levels of BDNF, phosphorylated tyrosine receptor kinase B (pTrkB), and phosphorylated CREB (pCREB) in the hippocampus and mPFC than those of control mice, whereas the expression of total β-actin, TrkB, and CREB among all groups remained unchanged. Similarly, Figure 2E reveals that the mice infused with SIK1-shRNA had notably fewer cells labelled with doublecortin (DCX) and fewer cells co-labelled with NeuN and BrdU in the dentate gyrus (DG) than those of control mice, representing a decrease in hippocampal neurogenesis due to SIK1 knockdown in the PVN. In contrast, infusion of Control-shRNA produced no influence on any of these biological indices. Taken together, these findings suggest that downregulated SIK1 expression in the PVN contributes to the pathogenesis of depression.
In addition, AAV-mediated selective overexpression of CRTC1 in the PVN was also adopted in this study, and the efficacy of AAV-CRTC1-EGFP is shown in Figure S3A. AAV-CRTC1 or AAV-Control was infused into the PVN of naïve C57BL/6J mice. After 14 d, it was found that compared with mice in the control group, the mice infused with AAV-CRTC1 but not AAV-Control exhibited evidently more immobility in the FST and TST as well as notably less sucrose preference and social interaction (Figure S3B). Meanwhile, CRTC1 overexpression in the PVN did not affect the locomotor activity of mice, as revealed by the OFT results (Figure S3B). Moreover, treatment with AAV-CRTC1 but not AAV-Control robustly promoted the plasma levels of corticosterone and ACTH in mice (Figure S3B). All these findings are consistent with the above SIK1-shRNA results. Therefore, enhanced CRTC1 expression in the PVN also contributes to depression.
The CRTC1-CREB-CRH pathway mediates the pro-depressant actions induced by SIK1 knockdown in the PVN
To understand whether the pro-depressant actions induced by SIK1 knockdown in the PVN required CRTC1, AAV-CRTC1-shRNA-EGFP was generated, and its efficacy was confirmed, as shown in Figure S4A. CRTC1-shRNA was infused into the PVN of naïve C57BL/6J mice, and after 14 d, SIK1-shRNA was also infused. After another 14 d, behavioural tests were performed. As shown in Figure S4B, compared with the control group, knockdown of CRTC1 in the PVN alone did not influence mouse behaviours in the FST, TST, sucrose preference test or social interaction test. However, CRTC1-shRNA pre-infusion significantly attenuated not only the tendency of SIK1-shRNA infusion to promote immobility in mice in the FST and TST but also the tendency of SIK1-shRNA infusion to reduce sucrose preference and social interaction in mice (Figure S4B). Moreover, CRTC1-shRNA pre-infusion notably prevented the enhancing effects of SIK1-shRNA infusion on the levels of plasma corticosterone and ACTH in mice (Figure S4B). The usage of Control-shRNA had no influence on these indices. In addition, Figure S4C reveals that the silencing effects of SIK1-shRNA and CRTC1-shRNA did not interfere with each other.
Then, to investigate whether the pro-depressant actions induced by SIK1 knockdown in the PVN required CREB, AAV-CREB-shRNA-EGFP was adopted, and its efficacy was confirmed, as shown in Figure S5A. By using an experimental procedure the same as described above, naïve C57BL/6J mice received infusions of CREB-shRNA (first) and SIK1-shRNA (second) into the PVN, followed by behavioral tests. The behavioural results are summarized in Figure S5B. Compared with the control group, knockdown of CREB in the PVN alone did not influence mouse behaviours. However, CREB-shRNA pre-infusion significantly prevented the pro-depressant effects of SIK1-shRNA infusion on mouse behaviours in the FST, TST, sucrose preference test and social interaction test. Moreover, CREB-shRNA pre-infusion fully abolished the promoting actions of SIK1-shRNA infusion on HPA activity, as revealed by the ELISA results involving plasma corticosterone and ACTH (Figure S5B). In addition, Figure S5C shows that the silencing effects of SIK1-shRNA and CREB-shRNA did not interfere with each other.
Furthermore, to explore whether the pro-depressant actions induced by SIK1 knockdown in the PVN required CRH, AAV-CRH-shRNA-EGFP was used, and its efficacy was confirmed, as shown in Figure S6A. As described above, CRH-shRNA (first) and SIK1-shRNA (second) were infused into the PVN of naïve C57BL/6J mice. The usage of CRH-shRNA significantly prevented the depression-like behaviours induced by SIK1-shRNA infusion, as revealed by the FST, TST, sucrose preference test and social interaction test (Figure S6B). Our ELISA results show that the usage of CRH-shRNA also notably blocked the promoting actions of SIK1-shRNA infusion on HPA activity (Figure S6B). Knockdown of CRH in the PVN alone had no influence. Figure S6C indicates that the silencing effects of SIK1-shRNA and CRH-shRNA did not interfere with each other.
Collectively, these findings suggest that the pro-depressant actions induced by SIK1 knockdown in the PVN require the CRTC1-CREB-CRH pathway.
Both genetic overexpression of SIK1 and genetic knockdown of CRTC1 in the PVN produced antidepressant-like effects in mice
We were interested in testing whether averting chronic stress–induced SIK1 downregulation in the PVN prevents depression. To achieve this purpose, AAV-mediated selective overexpression of SIK1 in the PVN was adopted, and the efficacy of AAV-SIK1-EGFP is demonstrated in Figure 4A. The CSDS-exposed mice were subjected to PVN infusion of AAV-SIK1 or AAV-Control, and after 14 d, behavioural tests were performed. As shown in Figure 4B, the CSDS-exposed mice treated with AAV-SIK1 displayed significantly decreased immobility in the FST and TST, increased sucrose preference and social interaction, and lower levels of plasma corticosterone and ACTH than the CSDS-exposed mice and the CSDS-exposed mice treated with AAV-Control. In parallel, the CUMS-exposed mice received PVN infusion of AAV-SIK1 or AAV-Control, followed by behavioural tests. As shown in Figure 4C, the CUMS-exposed mice treated with AAV-SIK1 also exhibited evidently less immobility in the FST and TST, higher sucrose preference, and reduced levels of plasma corticosterone and ACTH than the CUMS-exposed mice and the CUMS-exposed mice treated with AAV-Control. Treatment with AAV-Control did not influence these indices.
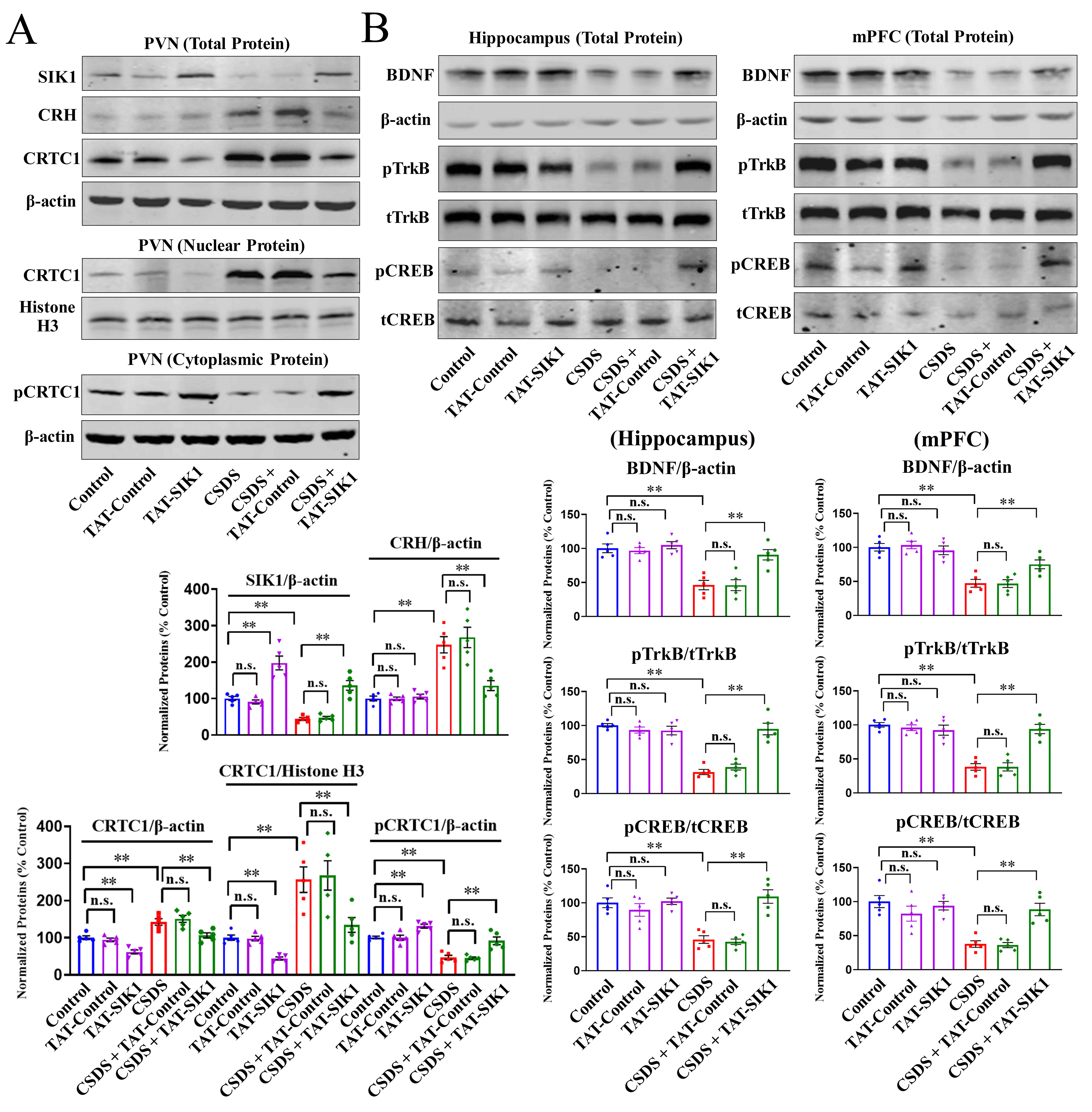
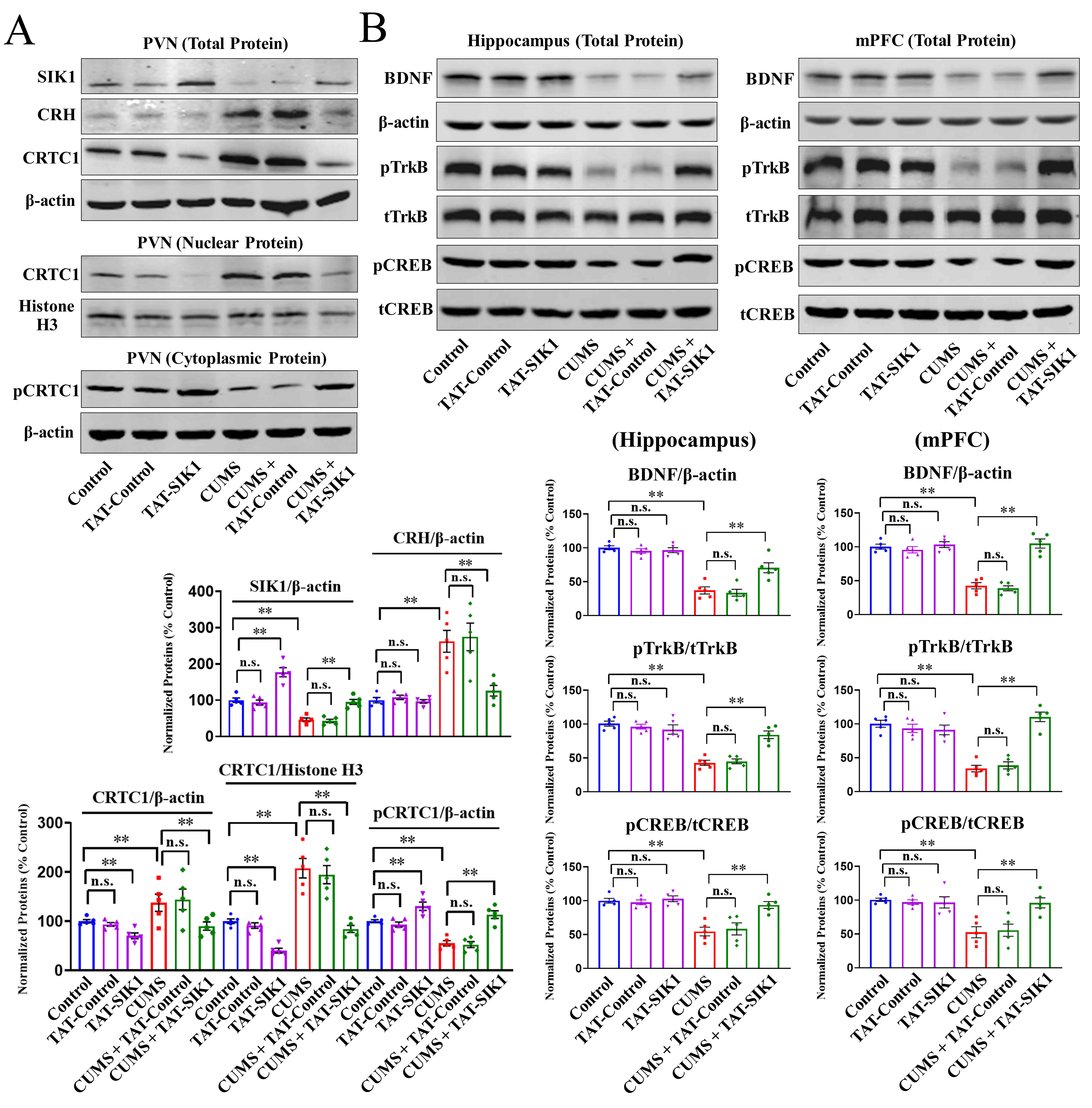
Then, changes in SIK1-CRTC1 signalling in PVN neurons between all groups were detected by western blotting, Co-IP, and immunofluorescence. AAV-SIK1 treatment not only fully reversed the CSDS-induced decrease in SIK1 (Figure 5A) and cytoplasmic pCRTC1 (Figure 5A) expression but also significantly prevented the CSDS-induced increase in the levels of total CRTC1 (Figure 5A and 6), nuclear CRTC1 (Figure 5A and 6), and CRTC1-CREB binding (Figure S13A). More importantly, AAV-SIK1 treatment notably antagonized the promoting effects of CSDS on CRH expression in PVN neurons (Figure 5A), consistent with the corticosterone and ACTH results in Figure 4B. Similarly, AAV-SIK1 treatment also fully reversed the CUMS-induced effects on SIK1 (Figure 5B), CRH (Figure 5B), nuclear CRTC1 (Figure 5B and 7), cytoplasmic pCRTC1 (Figure 5B), total CRTC1 (Figure 5B and 7), and CRTC1-CREB binding (Figure S13B) in PVN neurons. In addition, AAV-SIK1 treatment enhanced the expression of cytoplasmic pCRTC1 (Figure 5A and 5B) and downregulated the levels of total CRTC1 (Figure 5A, 5B, 6, and 7), nuclear CRTC1 (Figure 5A, 5B, 6, and 7), and CRTC1-CREB binding (Figure S13A and S13B) in the PVN of naïve control mice. Treatment with AAV-Control did not affect SIK1-CRTC1 signalling.
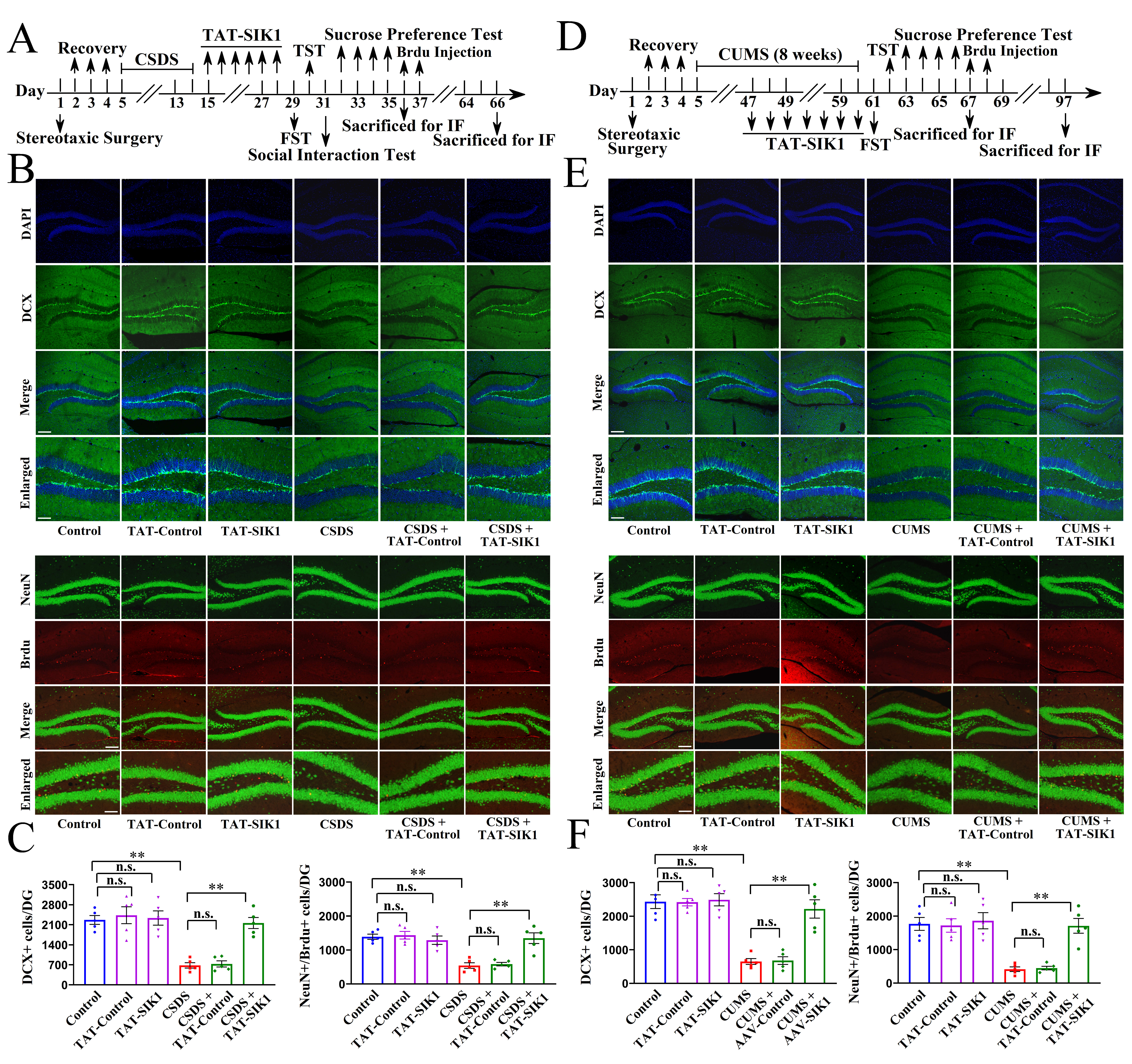
Next, the levels of the BDNF signalling cascade and neurogenesis among all groups were examined. AAV-SIK1 treatment evidently blocked both the CSDS-induced and CUMS-induced decreases in the expression of BDNF (Figure 5C and 5D), pTrkB (Figure 5C and 5D), and pCREB (Figure 5C and 5D) in the hippocampus and mPFC, whereas the expression of total β-actin, TrkB, and CREB in the two regions did not change. Similarly, AAV-SIK1 treatment markedly abolished both the CSDS-induced and CUMS-induced decrease in hippocampal neurogenesis, as revealed by Figure 8A and 8B. In addition, AAV-SIK1 treatment did not affect the levels of BDNF, pTrkB, pCREB, or neurogenesis in naïve control mice (Figure 5C, 5D, 8A, and 8B). Treatment with AAV-Control also produced no effects.
Furthermore, AAV-CRTC1-shRNA-EGFP was applied again. The CSDS-exposed mice were subjected to PVN infusion of CRTC1-shRNA or Control-shRNA, and after 14 d, behavioural tests were performed. As shown in Figure S7A, the CSDS-exposed mice treated with CRTC1-shRNA displayed significantly decreased immobility in the FST and TST, increased sucrose preference and social interaction, and lower levels of plasma corticosterone and ACTH than the CSDS-exposed mice and the CSDS-exposed mice treated with Control-shRNA. In parallel, the CUMS-exposed mice received PVN infusion of CRTC1-shRNA or Control-shRNA, followed by behavioural tests. As shown in Figure S7B, the CUMS-exposed mice treated with CRTC1-shRNA also exhibited evidently less immobility in the FST and TST, higher sucrose preference, and reduced levels of plasma corticosterone and ACTH than the CUMS-exposed mice and the CUMS-exposed mice treated with Control-shRNA. The usage of Control-shRNA had no influence.
Collectively, both genetic overexpression of SIK1 and genetic knockdown of CRTC1 in PVN neurons produced beneficial effects against chronic stress.
Stereotactic infusion of TAT-SIK1 into the PVN produced antidepressant-like effects in mice
Beyond genetic manipulation, we employed the TAT-SIK1 fusion protein (TAT-SIK1), a cell permeable peptide that can be efficiently introduced into neurons to increase the biological function of SIK1. The efficacy of TAT-SIK1 is demonstrated in Figure S8A. The CSDS-exposed mice were subjected to daily PVN infusion of TAT-SIK1 or TAT-Control (nonactive TAT-SIK1) for 14 d, followed by behavioural tests. As shown in Figure S8B, the CSDS-exposed mice treated with TAT-SIK1 displayed significantly decreased immobility in the FST and TST, increased sucrose preference and social interaction, and lower levels of plasma corticosterone and ACTH than the CSDS-exposed mice and the CSDS-exposed mice treated with TAT-Control. In parallel, the CUMS-exposed mice received daily PVN infusion of TAT-SIK1 or TAT-Control during the last 2 weeks, followed by behavioural tests. As shown in Figure S8C, the CUMS-exposed mice treated with TAT-SIK1 also exhibited evidently less immobility in the FST and TST, higher sucrose preference, and reduced levels of plasma corticosterone and ACTH than the CUMS-exposed mice and the CUMS-exposed mice treated with TAT-Control. Treatment with TAT-Control did not influence these indices.
Subsequently, changes in SIK1-CRTC1 signalling in PVN neurons among all groups were examined by western blotting, Co-IP, and immunofluorescence. TAT-SIK1 treatment not only fully reversed the CSDS-induced decrease in SIK1 (Figure S9A) and cytoplasmic pCRTC1 (Figure S9A) expression but also significantly prevented the CSDS-induced increase in the levels of total CRTC1 (Figure S9A and S11), nuclear CRTC1 (Figure S9A and S11), and CRTC1-CREB binding (Figure S13C). More importantly, TAT-SIK1 treatment notably antagonized the promoting effects of CSDS on CRH expression in PVN neurons (Figure S9A), consistent with the corticosterone and ACTH results in Figure S8B. Similarly, TAT-SIK1 treatment also fully reversed the CUMS-induced effects on SIK1 (Figure S10A), CRH (Figure S10A), nuclear CRTC1 (Figure S10A and S12), cytoplasmic pCRTC1 (Figure S10A), total CRTC1 (Figure S10A and S12), and CRTC1-CREB binding (Figure S13D) in PVN neurons. In addition, TAT-SIK1 treatment enhanced the expression of cytoplasmic pCRTC1 (Figure S9A and S10A) and downregulated the levels of total CRTC1 (Figure S9A, S10A, S11, and S12), nuclear CRTC1 (Figure S9A, S10A, S11, and S12), and CRTC1-CREB binding (Figure S13C and S13D) in the PVN of naïve control mice, further confirming the biological actions of TAT-SIK1. Treatment with TAT-Control did not affect SIK1-CRTC1 signalling.
Afterwards, the levels of the BDNF signalling cascade and neurogenesis in each group were assayed. TAT-SIK1 treatment evidently blocked both the CSDS-induced and CUMS-induced decreases in the expression of BDNF (Figure S9B and S10B), pTrkB (Figure S9B and S10B), and pCREB (Figure S9B and S10B) in the hippocampus and mPFC, whereas the expression of total β-actin, TrkB, and CREB in the two regions did not change. Similarly, TAT-SIK1 treatment markedly abolished both the CSDS-induced and CUMS-induced decreases in hippocampal neurogenesis, as revealed by Figure S14A and S14B. In addition, TAT-SIK1 treatment did not affect the levels of BDNF, pTrkB, pCREB, or neurogenesis in naïve control mice (Figure S9B, S10B, S14A, and S14B). Treatment with TAT-Control also produced no effects.
Combined with the above results involving AAV-SIK1, it can be concluded that SIK1 in the PVN is a feasible and novel antidepressant target.
Fluoxetine, paroxetine, venlafaxine, and duloxetine can modulate SIK1-CRTC1 signalling in PVN neurons
Currently, SSRIs and SNRIs are the most commonly used antidepressants in clinical practice. Fluoxetine and paroxetine are representative drugs of SSRIs. Venlafaxine and duloxetine are representative SNRI drugs. Here, we studied whether the antidepressant actions of fluoxetine, paroxetine, venlafaxine, and duloxetine involve the SIK1-CRTC1 system in the PVN.
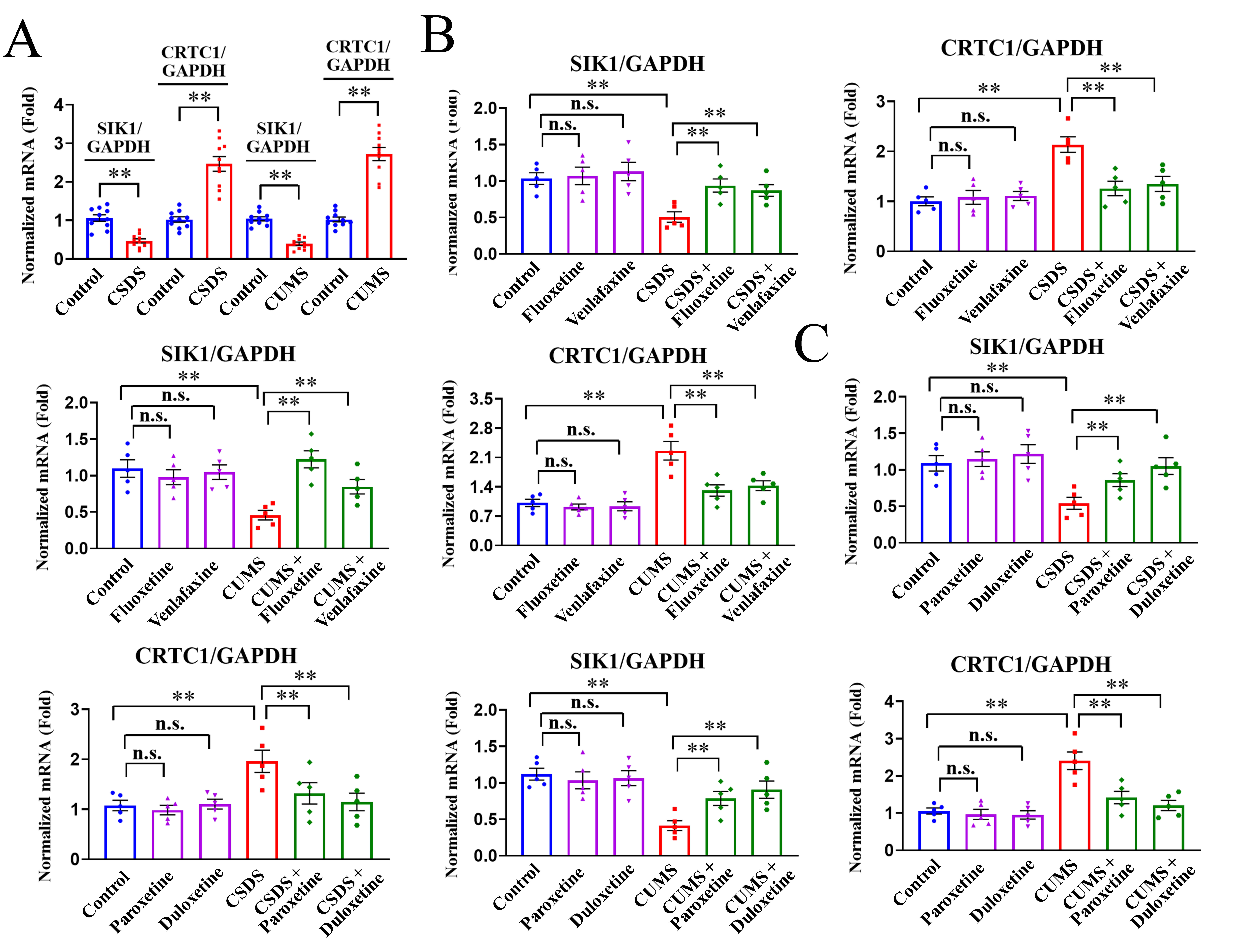
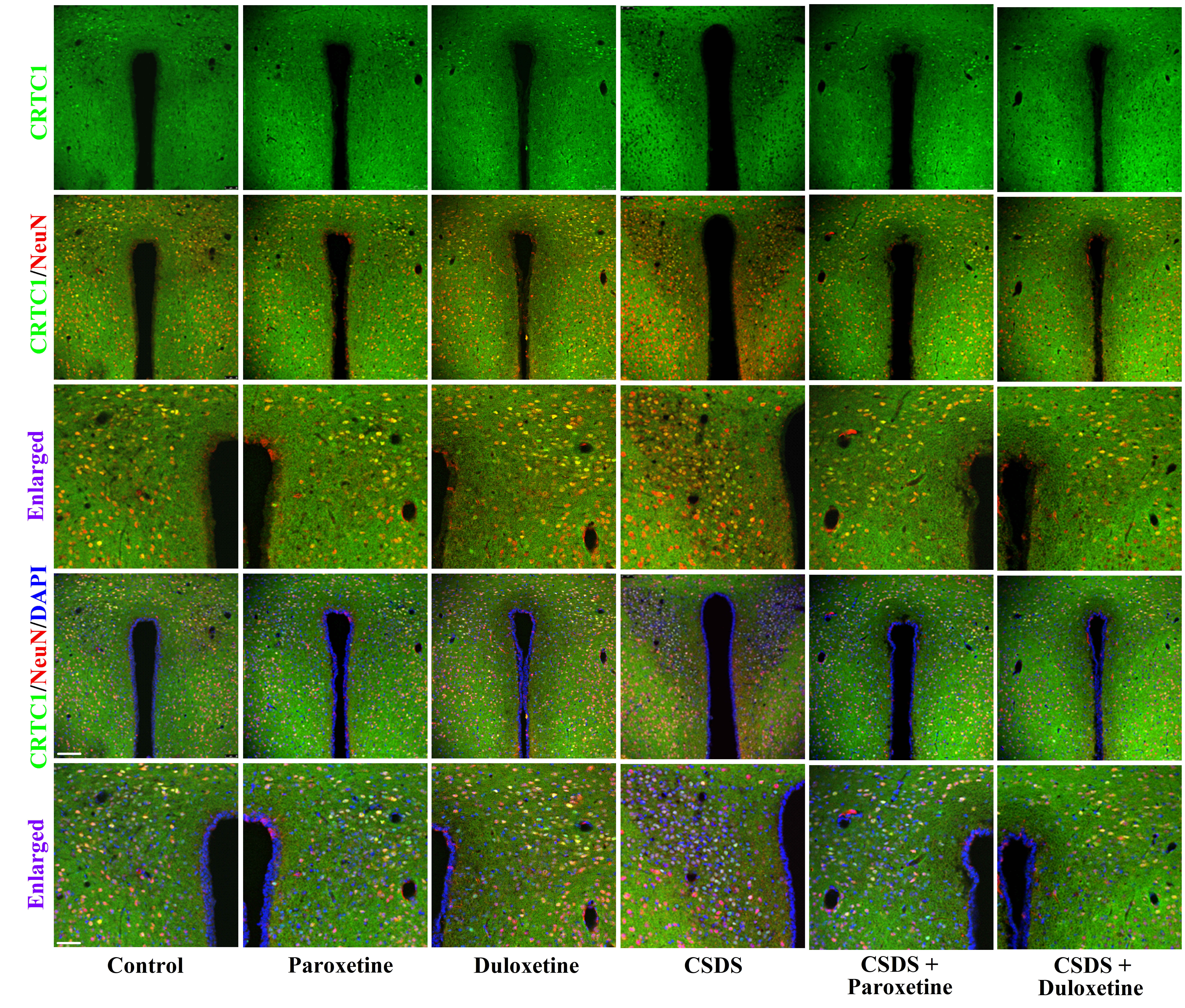
The CSDS-exposed mice received a daily injection of fluoxetine, paroxetine, venlafaxine, or duloxetine for 14 d, and behavioural tests were performed thereafter. As shown in Figures 9A and S15A, repeated treatment with the four drugs not only significantly reversed the CSDS-induced depression-like behaviours in mice but also entirely prevented CSDS from increasing the levels of plasma corticosterone and ACTH in mice. Then, it was found that all these drugs thoroughly reversed the CSDS-induced downregulation of SIK1 and upregulation of CRH in the PVN of mice (Figure 9C and S15B). Moreover, administration of the four drugs notably blocked not only the tendency of CSDS to promote nuclear CRTC1 (Figure 9C, 10, S15B, and S17), total CRTC1 (Figure 9C, 10, S15B, and S17), and CRTC1-CREB binding (Figure S19A and S19C) but also the inhibitory effects of CSDS on cytoplasmic pCRTC1 in the PVN (Figure 9C and S15B) of mice. Furthermore, administration of the four drugs evidently increased the SIK1 mRNA level and decreased the CRTC1 mRNA level in the PVN of the CSDS-exposed mice (Figure S1B and S1C). In addition, none of these drugs affected the SIK1-CRTC1 system in the PVN of naïve control mice (Figure S1B, S1C, 9C, 10, S15B, and S17).
The CUMS-exposed mice were subjected to daily injection of fluoxetine, paroxetine, venlafaxine, or duloxetine during the last 2 weeks, and then, behavioural tests were performed. Figures 9B and S16A show that the four drugs also demonstrated notable antidepressant actions in the CUMS model. More importantly, in parallel with the CSDS results, the CUMS-induced effects on the expression of SIK1 (Figure 9D and S16B), CRH (Figure 9D and S16B), nuclear CRTC1 (Figure 9D, 11, S16B, and S18), total CRTC1 (Figure 9D, 11, S16B, and S18), cytoplasmic pCRTC1 (Figure 9D and S16B), and CRTC1-CREB binding (Figure S19B and S19D) in the PVN of mice were all thoroughly reversed by administration of these drugs, as revealed by western blotting, Co-IP, and immunofluorescence. In addition, treatment with these drugs all significantly prevented the effects of CUMS on SIK1 mRNA and CRTC1 mRNA in the PVN of mice (Figure S1B and S1C).
In summary, fluoxetine, paroxetine, venlafaxine, and duloxetine can modulate SIK1-CRTC1 signalling in PVN neurons.
The SIK1-CRTC1 system in the PVN is necessary for the antidepressant actions of fluoxetine, paroxetine, venlafaxine, and duloxetine
AAV-SIK1-shRNA-EGFP was further used to determine whether SIK1-CRTC1 signalling in PVN neurons participates in the antidepressant mechanisms of fluoxetine, paroxetine, venlafaxine, and duloxetine.
C57BL/6J mice pre-infused with SIK1-shRNA were subjected to 10 d of CSDS and then daily injection of fluoxetine, paroxetine, venlafaxine, or duloxetine for 14 d, followed by behavioural tests. As shown in Figures 12A and S20A, knockdown of SIK1 in the PVN significantly attenuated the reducing effects of all these drugs on the immobility duration of the CSDS-exposed mice in the FST and TST. Additionally, knockdown of SIK1 in the PVN also abolished the promoting effects of the four drugs on the sucrose preference and social interaction of the CSDS-exposed mice. Moreover, C57BL/6J mice pre-treated with SIK1-shRNA were exposed to 8 weeks of CUMS and daily treatment with fluoxetine, paroxetine, venlafaxine, or duloxetine during the last 2 weeks, and behavioural tests were then performed. Pre-treatment with SIK1-shRNA notably antagonized the antidepressant effects of all these drugs against CUMS in the FST, TST and sucrose preference test (Figure 12B and S20B). In contrast, the usage of Control-shRNA produced no effects on the behavioural results of mice (Figure 12A, 12B, S20A, and S20B). Combined with the above results, the SIK1-CRTC1 system in the PVN is required for the therapeutic actions of the four antidepressants used in clinical practice.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}