Effects of N deficiency and re-supply treatments on lipid peroxidation and ROS accumulation in tomato roots
Membrane lipid peroxidation often occurs when plant organs are aged or damaged under adversity. MDA is the final decomposition product of membrane lipid peroxidation, and its content can reflect the degree of damage to plants under adversity. In this experiment, the MDA content of samples was measured at four treatment time points: CK, CK-N, T1 and T2. After two days of N deficiency treatment, the MDA content increased by 89.90 % compared with that of normal tomato seedlings (Fig. 1a). Compared with the control group, MDA content still increased by 38.57 % after re-supplying N for 6 h, and basically returned to normal level after re-supplying N for 24 h.
The ROS content in tomato roots was labeled with fluorescent probe H2DCF-DA (Fig. 1b). The fluorescence degree of ROS in roots treated with N deficiency was increased dramatically, which indicated that the oxidative damage caused by N deficiency was aggravated. The fluorescence intensity decreased after re-supplying N, which indicated that the oxidative damage was relieved to some extent after re-supplying N.
Effects of N deficiency and re-supply treatments on antioxidant enzyme activities and ratio of AsA/DHA and GSH/GSSG in tomato roots
As shown in Fig. 2, the activities of SOD, CAT, APX, and MDHAR in tomato roots were all decreased after N deficiency stress compared with the control. SOD activity decreased by 38.02 %, 22.01 % and 19.71 % after 2 days of N deficiency, N re-supply for 6 h and 24 h (Fig. 2a). CAT activity decreased significantly by 48.50 % after N deficiency stress, and continued to decrease to 59.72 % after 6 h of N re-supply (Fig. 2b). APX is an antioxidant enzyme that mainly catalyzes AsA to capture H2O2 in chloroplasts and cytoplasm of plant cells. The APX activity in the roots of tomato seedlings decreased by 32.78 % after N deficiency, and continued to decrease to 43.45 % after 6 h of N re-supply and then increased by 25.02 % after 24 h of N re-supply (Fig. 2c). MDHAR is an important enzyme to regenerate AsA in AsA-GSH cycle. MDHAR activities decreased by 60.13 % after N deficiency and then increased after N re-supply (Fig. 2d).
The ratio of ASA/DHA and GSH/GSSG in tomato seedling roots decreased after N deficiency and then increased after N re-supply (Fig. 2e, f). Compared with the control group, the ratio of GSH/GSSG and ASA/DHA decreased by 29.15 % and 36.22 %, respectively, after 2 days of N deficiency. The ratio of ASA/DHA increased by 16.15 %, 21.59 % and the ratio of GSH/GSSG increased by 8.93 %, and 17.18 %, respectively, after 6 h and 24 h of N re-supply.
Effects of N deficiency and N re-supply treatments on NO3−contents, nitrate reductase activity in tomato roots
After N deficiency for 2 days, the NO3− content decreased by 39.61 %, compared with the control (Fig. 3a). The NO3− content in tomato roots increased by 17.56 % after re-supplying N for 6 h. The N re-supply for 24 h treatments basically restored NO3− accumulation to normal levels.
Nitrate reductase (NR) is a key enzyme in the plant nitrate-assimilation process. Therefore, effects of N deficiency and the N re-supplying treatment on the NR activity in tomato roots were investigated. As shown in Fig. 3b, N deficiency treatments significantly reduced the NR activity by 54.78 % in tomato roots. However, the inhibitory effect under conditions of N deficiency was relieved by the N re-supply treatment.
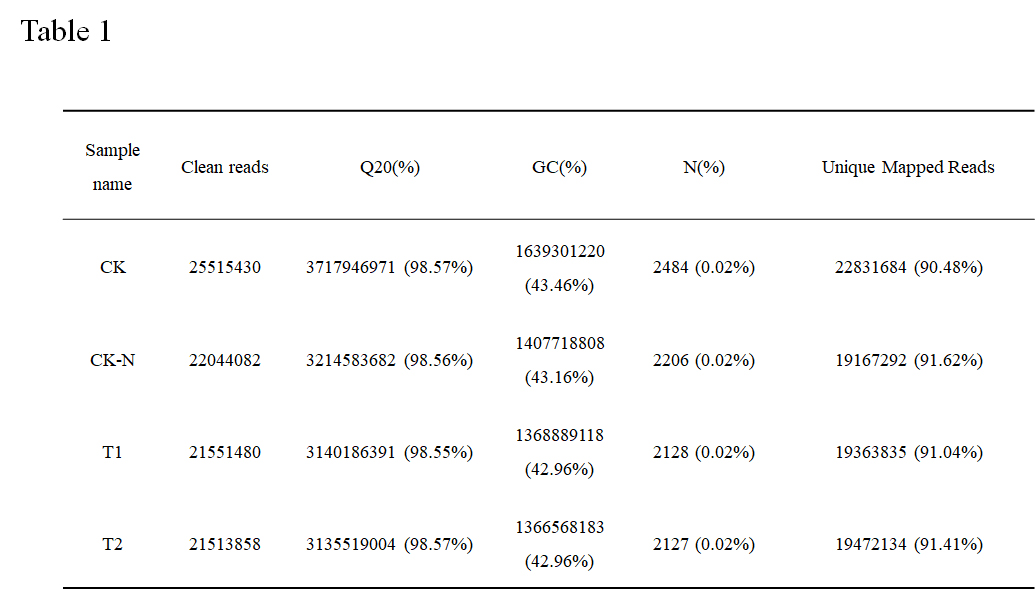
Quality analysis of sequencing results
To investigate the molecular response of N starvation and re-supply to tomato seedling, RNA-seq technology was used. The sample of tomato roots of the control and the treatment group were sequenced by Illumina HiSeqTM. In order to ensure the quality of the data, the original data should be quality-controlled before information analysis. Removed reads with adapter, reads with N ratio greater than 10 %, and low-quality reads, CK, CK-N, T1, and T2 have 25515430, 22044082, 21551480, and 21513858 clean reads, and the percentage of Q20 (the quality of the bases after filtering is not less than 20 Proportion) reached 98.32 %, 98.35 %, 98.27 %, and 98.32 %, all of which are greater than 90%. The quality is qualified and meets the requirements of subsequent analysis. In this study, tomato samples of treatments CK, CK-N, T1, and T2 treatments had GC contents of 43.46 %, 43.16 %, 42.96 %, and 42.96 %. Removed reads on the ribosome, four processed Unmapped Reads were got, and the total reads are 25233998, 20919716, 21269156, and 21301956, respectively. Number of reads on the reference genome for unique alignment and the proportion of the total (Unique Mapped Reads) alignment are 22831684 (90.48 %), 19167292 (91.62 %), 19363835 (91.04 %) and 19472134 (91.41 %). The above data will be used for assembly and analysis of the transcriptome (Table 1).
PCA results showed that there was a significant difference in gene expression between the CK group and the CK-N group. The gene expression of the T1 group showed a recovery phenomenon, and the gene expression of the CK group samples was similar to that of the T2 group samples (Fig. S1). The results showed that the gene expression level basically recovered to that of the CK group of the hydroponic tomato roots resumed N culture for 24 h after 2 days of N deficiency.
Analysis of differences between samples
The R-based software package edgeR was used to process the RNA-seq data for pairwise samples or between groups with significant differences. FDR and log2FC were used to screen for differential genes, and the screening conditions were FDR < 0.05 and | log2FC | > 1. As shown in Fig. 4a, compared the CK with CK-N, there were 4010 significantly different genes, of which 1766 were significantly up-regulated and 2244 were significantly down-regulated. Comparing the CK-N and T1 groups, there were 5441 significantly different genes, of which 3534 were significantly up-regulated and 1907 were significantly down-regulated. Comparing the CK-N and T2 groups, there were 4235 significantly different genes, of which 2233 were significantly up-regulated and 2002 were significantly down-regulated. Compared with the T1 and T2 groups, there were 2817 significantly different genes, of which 839 were significantly up-regulated and 1978 were significantly down-regulated.
To identify common and unique DEGs in response to N starvation and re-supply treatments, venn graphs were plotted. We analyzed DEGs that were transcriptionally regulated at different treatment, and a total of 430 common DEGs were identified among the four libraries (Fig. 4b).
GO function annotation analysis of different expressed genes
The number of genes with significant differences in GO function enrichment between each 2 groups is shown in Table S2. These differentially expressed genes are annotated into three directions: biological processes, cell components and molecular functions, involving a total of 45 functional entries. The GO classification charts of Ck-N-vs-T1, Ck-N-vs-T2 and T1-vs-T2 are shown in Fig S2, S3, S4. The GO classification map of CK and CK-N are shown in Fig. 5. In biological processes, differentially expressed genes are most abundant in cellular processes, metabolic processes and single-organism processes. In cell components, differentially expressed genes are mainly concentrated in cell, cell part and membrance. In terms of molecular function, differentially expressed genes are mainly enriched in catalytic activity and binding function items.
Differential gene KEGG Pathway analysis
In order to further understand the biological function of genes and determine the most important biochemical metabolic pathways and signal transduction pathways involved in DEGs, we performed pathway significant enrichment analysis. 7135 differentially expressed genes were annotated in the KEGG database. In CK and CK-N, CK-N and T1, CK-N and T2, T1 and T2, they were annotated to 124, 126, 125, and 113 pathways, respectively. The top 20 pathways with abundant genes are shown in Fig. 6 and Fig. S5, S6, S7. Among them, the differentially expressed genes are mainly involved in phenylpropanoid biosynthesis, amino sugar and nucleotide sugar metabolism, cysteine and methionine metabolism, starch and sucrose metabolism, N metabolism and other pathways.
Differential expression of N metabolism genes in tomato
The metabolic process of N in plants includes complex mechanisms such as absorption and transport, assimilation and reuse. As shown in Fig. 7, differentially expressed genes were found in this N metabolism pathway. Compared with the control group, 7 DEGs expressions were down-regulated and 1 DEGs expression was up-regulated in hydroponic tomato roots after N deficiency. The down-regulated genes were nitrate transporter (Nrt, Solyc06g010250.3), nitrate reductase (NR, Solyc11g013810.2), nitrite reductase (NiR, 1.7.7.1, Solyc01g108630.3), alpha carbonic anhydrase (14.2.1.1, Solyc09g009830.3). Besides, Beta carbonic anhydrase 5 (14.2.1.1, Solyc09g010970.3), Glutamate dehydrogenase (GDH, 1.4.1.3, Solyc05g052100.3), glutamine synthetase (GS, 6.3.1.2, Solyc01g080280.3). Glutamate synthase 1 (GOGAT, 11.4.1.13 and 1.4.1.14 have the same Nr annotation of Solyc03g083440.3) have up-regulation expression.
The genes that are down-regulated or up-regulated due to N deficiency stress gradually returned to normal expression levels after 1 day of N restoration.
The expression of antioxidant enzyme genes in qRT-PCR and RNA-seq analysis
qRT-PCR analysis was performed on the transcription level of SlSOD, SlCAT, SlAPX and SlMDHAR (Fig. 8). After 2 days of N deficiency in tomato seedlings, the relative expression of SlSOD, SlCAT, SlAPX and SlMDHAR genes in tomato roots decreased by 97.37 %, 94.86 %, 73.83 % and 83.45 %, respectively, compared with the control group. The relative expression of SlSOD and SlCAT genes increased by 6.55 and 3.76 times respectively after N re-supply for 6 h. The relative expression of SlAPX and SlMDHAR genes decreased by 74.63 % and 51.24 % respectively after N re-supply for 6 h. The relative expression of SlSOD, SlCAT, and SlMDHAR genes increased by 7.54, 8.53 and 1.91 times, and the relative expression of SlAPX genes decreased by 43.89 %, after N re-supply for 24 h.
The mRNA expression of SlTrxh, SlPrx and SlGrx was also analyzed by qRT-PCR in the roots of tomato seedling after the N deficiency and re-supply. SlTrxh, SlPrx and SlGrx expression decreased by 56.73 %, 93.27 % and 87.31 %, respectively, compared with the control group after 2 days of N deficiency. SlTrxh expression decreased by 53.74 % compared with the control group after 6 h of N re-supply. SlPrx and SlGrx expression increased by 4.77 and 1.03 times of the control group after 6 h of N re-supply. The mRNA expression of SlPrx and SlGrx increased significantly after 24 h of N re-supply.
We analyzed the transcriptome, and the results are basically consistent with the real-time PCR data (Table S3), indicating that the transcriptome data is reliable.
The expression of N metabolism genes in qRT-PCR and RNA-seq analysis
Compared with the control group, the transcription levels of SlNRT2.4, SlNR, SlNiR and SlGS in hydroponic tomato seedlings were down-regulated by 65.43%, 13.22%, 73.11% and 54.72%, respectively, after N starvation (Fig. 9). Compared with the control group, the transcription level was up-regulated by 1.17, 2.24, 2.15 and 0.59 times after N re-supplying for 24 h. Compared with the control group, the transcription level of SlGOGAT gene after N starvation treatment was increased by 2.19 times, and after 24 h of N re-supplying, the transcription level was only 1.07 times that of the control group, which basically recovered to the level of the control group.
{kind=link}