Transcripts encoding neurodevelopmental regulators are m6A modified in neuroblasts and neurons
Near the end of Drosophila larval neurogenesis, the combined brain lobes contain approximately 10,000 neurons, roughly 500 glia, and only 200 neuroblasts [21, 22]. To increase representation of the neuroblast methyltranscriptome, we used a genetic modification that causes neuroblasts to undergo symmetric self-renewing divisions, thus generating larval brains with abundant ectopic neuroblasts and relatively few neurons [21]. In these experiments, we used insc-Gal4 to drive expression of UAS-aPKCCAAX in neuroblasts and harvested larval brains at 96–102 hours after larval hatching (ALH) as a source of “neuroblast-biased RNA”. In contrast, we used wildtype larval brains at 96–102 hours ALH as a source of “neuron-biased RNA” since neurons are vastly more abundant than any other cell type at this stage. In addition to collecting RNA samples that cover the neuron and neuroblast methyltranscriptomes, we collected RNA from stage-matched brains of Mettl3 null larvae to obtain negative control “m6A null RNA”. Brain RNA from each genotype was split in two; half was used for quantification of total mRNA abundance (input RNA-seq) and half was used for methyltranscriptome purification using anti-m6A immunoprecipitation (meRIP-seq) [13]. This experimental design is summarized in Fig. 1A. As a first step, we used input RNA to test for differential abundance of known neuroblast or neuron-specific mRNAs in the neuron-biased and neuroblast-biased samples. We confirmed that insc-Gal4 > UAS-aPKCCAAX samples are enriched in neuroblast-specific transcripts and depleted of neuron-specific transcripts (Fig. 1B).
Subsequent meRIP-seq analysis of neuroblast-biased, neuron-biased and m6A-null RNA samples identified 867 m6A targets in the larval brain (Fig. 2A and SUPPLEMENTAL TABLE 1). 634 of these targets (73%) were also identified in adult Drosophila heads by Kan et al., revealing a high degree of m6A conservation across life cycle stages. As previously described, the m6A-null meRIP-seq data were useful for identifying “background” signal. This allowed high-confidence target identification and more accurate mapping of m6A peaks along a transcript: only peaks that were significantly enriched compared to m6A-null meRIP were included. Using this approach, we found that the vast majority of m6A peaks in the neuroblast-biased and neuron-biased transcriptomes map to the 5’ UTR (Fig. 2B). We used sequences from the combined neuron-biased and neuroblast-biased datasets to search for motifs associated with m6A and found significant enrichment of an AAACV motif. This motif contains the invariant AAAC core identified in other Drosophila m6A mapping studies [9] [7].
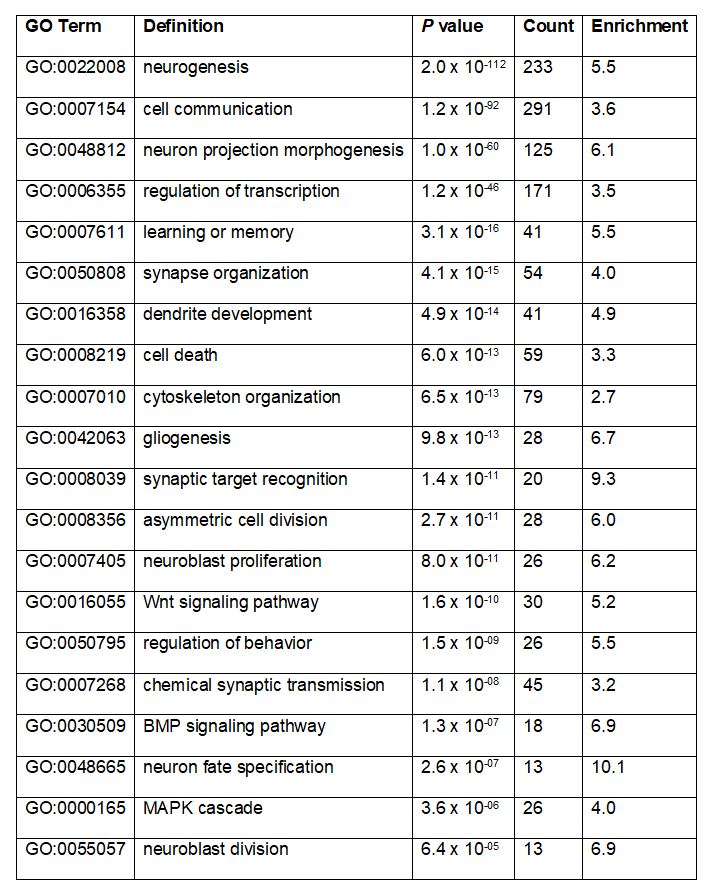
To gain insight into the potential roles of m6A in larval brain development, we used gene ontology analysis to identify functional categories overrepresented among m6A targets. This revealed significant enrichment of transcripts encoding regulators of essential neurodevelopmental processes, such as “synapse organization”, “dendrite development”, “neuroblast proliferation” and “neuron fate specification” in addition to processes known to be broadly important for development, such as “cell death”, “cytoskeleton organization”, and “Wnt signaling pathway” (Table 1). As expected, the combined profiling of neuroblast-biased and neuron-biased brains allowed identification of a large number of m6A targets (233 genes) that were not identified by previous m6A mapping in adult heads [9]. This novel set of m6A targets includes many genes known to regulate neuroblast proliferation, asymmetric cell division, neuron fate specification and axon pathfinding (Fig. 3A).
Comparing neuron-biased and neuroblast-biased meRIP-seq data revealed several genes with higher m6A peaks in one genotype or the other, potentially indicating cell type-specific differences in m6A stoichiometry (Fig. 3B). To test this possibility, we normalized meRIP-seq ratios (neuron-biased / neuroblast-biased) to input ratios (neuron-biased / neuroblast-biased). This identified cases where differential m6A peaks could be explained by differences in total transcript abundance. Following normalization for input reads and filtering for genes with statistically significant differences, we did not identify any evidence of differential m6A stoichiometry (Fig. 3C). 135 genes had approximately equal input expression levels (fold-change ≤ 1.5 and no statistically significant difference between neuroblast-biased and neuron-biased input mRNA abundance), but none of these “uniformly” expressed transcripts showed evidence of elevated m6A frequency in neuroblast-biased or neuron-biased brains. This suggests that elevated m6A peaks in neuroblast-biased brains, as shown for Sp1 and run in Fig. 3B, are due to elevated expression of the corresponding transcripts in neuroblasts. The converse is true for elevated m6A counts in neuron-biased brains. We further tested this conclusion using m6A immunoprecipitation and RT-qPCR of Sp1 and run (Fig. 3D). 5S rRNA served as a negative control in these experiments as it was not identified as a m6A target in our experiments and is known to lack methyladenosine in metazoans [23]. meRIP and RT-qPCR confirmed Sp1 and run as m6A targets and ruled out differential m6A between neuroblast-biased and neuron-biased brains. Overall, our m6A mapping indicates that m6A is selectively targeted to neurodevelopmental genes in neuroblasts and neurons and that for transcripts present in both cell types, the degree of m6A modification is largely constant.
M6a Correlates With Low Translation Efficiency And Low Mrna Stability
Given that m6A has been implicated in a range of mRNA metabolic processes, we next sought clues to the molecular function of m6A during larval brain development. Akhtar et al. identified a role for m6A and the nuclear m6A reader in enhancing transcription by relieving RNAP II pausing at target genes. This was demonstrated in Drosophila S2 cells and the phenomena has not been described in vivo or in a developmental context. To test this possible function, we used RNA-seq measurements of total mRNA abundance from wildtype brains and Mettl3 null brains. We reasoned that if m6A significantly enhances transcription in larval brains, the absence of m6A would result in decreased target abundance due to increased RNAP II pausing. As previously shown for adult Drosophila heads [9], this analysis failed to identify a strong directional relationship between m6A and transcript abundance (Fig. 4A). We also tested for a relationship between m6A and translation efficiency (TE). Using the adult head ribosome profiling data analyzed by Kan et al. [9], we found a similar significant enrichment of m6A in mRNAs with low translation efficiency (Fig. 4B).
Next, we tested for any relationship between m6A and mRNA stability. We obtained mRNA half-life measurements for neural progenitors and neurons using EC-tagging pulse-chase [14]. Briefly, this approach uses targeted expression of a cytosine deaminase-uracil phosphoribosyltransferase (CD:UPRT) fusion enzyme to convert 5-ethynylcytosine (EC) into 5-ethynyluridine (EU)-monophosphate in specific cell types. EU is incorporated into nascent mRNAs of target cells and the tagged RNAs can be purified after “pulse” feeding 5EC and at subsequent “chase” timepoints in which excess uridine is provided to ensure no new tagged transcripts are made. We used insc-Gal4 to express UAS-CD:UPRT in neural progenitors and nSyb-Gal4 to express UAS-CD:UPRT in neurons. Globally, neural progenitor and neuron transcriptomes had similar half-life distributions (Fig. 4C and SUPPLEMENTAL TABLE 2), indicating that transcriptome-wide mRNA decay kinetics do not significantly differ between neural progenitors and neurons. However, differences were revealed when we analyzed the half-lives of m6A targets: there was no relationship between m6A and mRNA stability in neurons (Fig. 4D), while m6A targets were significantly less stable in neuroblasts (Fig. 4E).
To further investigate the different relationships between m6A and stability in neuroblasts and neurons, we directly compared the half-lives of m6A targets in each cell type and found that 185 m6A targets are at least 1.5-fold more stable in neurons (Fig. 5A). If one assumes m6A directly affects mRNA stability, this differential decay is surprising given that our data suggest m6A is constant between neuroblasts and neurons. Differential stability could be caused by varied Ythdf expression, however; our EC-tagging data (data not shown) and prior transcriptome profiling of purified neuroblasts and neurons [24] show that Ythdf mRNA is present at equally high levels in progenitors and neurons. Alternatively, these data agree with a model in which the difference between neuroblasts and neurons is due to m6A-independent stabilization of target mRNAs in neurons. GO analysis of the neuron-stabilized m6A targets revealed enrichment of transcripts involved in neuron-specific functions such as “synapse assembly”, “dendrite development” and “axon guidance” (Fig. 5B), supporting the model that these transcripts are likely selectively stabilized to support the needs of mature neurons. We conclude that neuron-specific stabilization of m6A targets explains the lack of correlation between m6A and half-life in neurons.
M6a And Ythdf Enhance Target Protein Expression In Larval Brains
The analyses described above reveal correlations between m6A, mRNA translation and mRNA decay, but these findings do not reveal underlying mechanisms or causal relationships. With respect to translation, two mechanisms have been described in Drosophila: translation inhibition that requires Fmr1 [7] and Ythdf-dependent translation enhancement [9]. Comparing our m6A targets with previously identified m6A-dependent Fmr1 targets in the larval central nervous system revealed an overlap of only 5.8% (50 genes). Since the majority of our targets are not predicted to be regulated by Fmr1, we conclude that the translation enhancing effect may be more relevant. With respect to mRNA stability, 3’ UTR m6A in mammalian transcripts induces decay via DF proteins recruiting the CCR4-NOT deadenylase complex [5] but a decay pathway triggered by 5’UTR m6A has not been described in any species. Instead, we predict that the relationship between m6A and mRNA is indicative of a compensatory mechanism, similar to that described for translation efficiency. In this case, we predict that 5’UTR m6A enhances translation of low stability transcripts whose decay is regulated by m6A-independent mechanisms.
According to the translation enhancement model, Mettl3 deletion should decrease target protein production and Ythdf overexpression should increase target protein production. To test this model in the developing larval brain, we performed quantitative immunofluorescent imaging of proteins encoded by m6A targets in wildtype brains, Mettl3 null brains and Ythdf overexpressing brains (overexpressing Ythdf in neural progenitors using insc-Gal4 > UAS-Ythdf). We measured immunofluorescent signal for two m6A targets, the transcription factor Runt (Run) and the cell cycle regulator Cyclin D (CycD), in addition to one non-target, the transcription factor Asense (Ase). Translation efficiency data are not available for run and ase, likely because these genes are not expressed or are only expressed at low levels in adult brains, but the TE value for CycD in adult heads is 1.17 compared to an average value of 1.37 [25]. In contrast to the TE data, mRNA stability data are available for each of these genes. In neural progenitors run decays very rapidly (half-life of 5.1 minutes) and is more stable in neurons (half-life of 17.6 minutes). In larval brains, CycD and ase expression is primarily restricted to neural progenitors and we therefore only obtained progenitor-specific decay measurements for these transcripts: CycD has a half-life of 136.3 minutes and ase has a half-life of 16.1 minutes.
Runt expression in neuroblasts changed in a manner corresponding to the translation enhancement model: Runt signal decreased in Mettl3 null neuroblasts and increased in Ythdf overexpressing neuroblasts (Fig. 6A). In neurons, Runt signal was unaffected by loss of Mettl3 but increased in Ythdf overexpressing brains. Similar to Runt, CycD protein levels decreased in Mettl3 null neuroblasts, but Ythdf overexpression did not alter CycD abundance (Fig. 6B). Finally, as expected, neither Mettl3 loss-of-function or Ythdf overexpression altered the abundance of the non-target Asense (Fig. 6C). The run and CycD data support our prediction that m6A does not induce mRNA decay; if this were the case, Mettl3 deletion would most likely increase protein levels (we observe the opposite effect) and Ythdf overexpression would decrease protein levels (again, we see the opposite). Instead, these results support the model that 5’UTR m6A enhances translation of target mRNAs in the developing nervous system.
{kind=link}