This is the first study that widely surveyed the field strains of tropical bed bug in Iran via bioassay and molecular analysis. There has previously been no available information regarding in kdr and sequence data of vgsc gene of this species in studied areas. Therefore, generating relevant bioassay and genetic data were of high priority for investigations focusing on bed bug controlling in this country.
In view of the fact that we did not have a standard susceptible strain, as a limitation of this study, comparison of the bioassay data between studied populations revealed low levels of knockdown and mortality rates to permethrin in all of the field strains of tropical bed bug. Even though the knockdown and the mortality rates are not indicators of the same mechanisms of resistance and evolutionary selection, the correlation between these indexes is strong enough to identify permethrin resistance in the field populations. These values cannot be confidently related to the permethrin resistance phenotype and to identify the actual values further research is suggested with the provision of sensitive strain.
In this study, a similar phenomenon, resistance to insecticides, was found in the field strains of C. hemipterus to different pyrethroids (bifenthrin, cypermethrin, esfenvalerate, and etofenprox) from Thailand [41], DDT and deltamethrin residues from Malaysia [44], a mixture of deltamethrin and imidacloprid from Australia [51], and permethrin and deltamethrin from Sri Lanka [26]. These conditions were also represented in different populations of the common bed bug for pyrethroids from USA [23], permethrin and deltamethrin from Australia [52], allethrin from Sydney [43], imidacloprid and β-cyfluthrin from Malaysia [53, 54], several neonicotinoids (imidacloprid, acetamiprid, thiamethoxam, and dinotefuran) from USA [55], bifenthrin from Germany [56], a mixture of deltamethrin and imidacloprid from Australia [52], and bendiocarb from Paris [57]. Based on the bioassay results and the findings of previous studies [26, 41, 44, 51], all the bed bug populations in the present study manifested resistance to permethrin. Thus, the reconsideration of control methodologies is of paramount importance in the studied areas.
From the bioassay data, it was obtained that C. hemipterus had higher resistance to permethrin compared to the common bed bug, and KR values were found to be 5–7 in this bed bug. Due to the lack of a susceptible strain, the main limitation of this study, the estimated values of KR, using C. lectularius Kh field samples, cannot be a real value of resistance factor, as they are overestimated. Despite the absence of M918I and L1014F mutations in common bed bug samples, the possibility of V419L mutation [9, 19, 22–24], detoxification of enzymes (Dang, et al., 2017a), and heterogeneity of field samples could enhance the baseline susceptible data in this bed bug. Nevertheless, the higher value of KR, explaining the high frequency of kdr alleles, is strong evidence for the development of pyrethroid resistance. Therefore, the inclusion of resistance management in modified control programs is an important issue that needs to be taken into account.
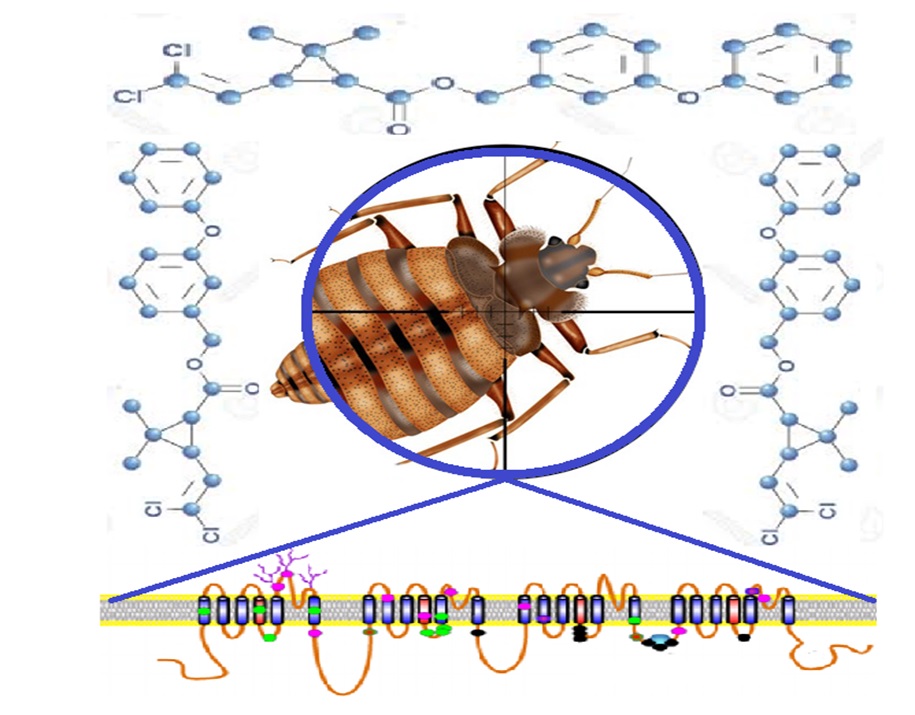
With the help of the SYBR green RT-PCR assay, the major kdr mutations were identified in all of the tropical bed bug samples. This finding, the same as the results of the previous studies [9, 25, 58, 59], confirms the presence of high levels of pyrethroid resistance among our studied populations. This is a fact that makes the reconsideration of control programs, necessary and can be used as baseline information for proper management of resistance in other areas. In the present study, the high sensitive/specific, less time-consuming, and low-cost PCR method calculated precisely the frequency of major kdr mutations. Therefore, it is suggested to identify the allelic frequency of the double-mutation, M918I and L1014F, in different populations of tropical bed bugs.
The TaqMan real-time PCR, a high throughput "closed-tub" diagnostic assay, developed in the current study detected the M918I allele in all of the C. hemipterus individuals without post PCR processing. This method revealed that all the samples of the studied populations carry the M918I mutation. Owing to the limitation of sensitive strains, the presence of M918 mutation in all specimens may seem to be overstated. Nonetheless, it is a fact that must be accepted because nucleotide sequencing of samples identified double mutations, M918I and L1014F, in all of the studied populations. The role of these mutations in kdr-type resistance, reduction of target site sensitivity to pyrethroids, has been proven within the tropical bed bug and a wide range of insects [9–13, 16, 25, 60, 61]. The association of M918I mutation with super-kdr resistance in C. hemipterus and several other insects has been affirmed in earlier studies [61]. From the result of the present investigation and those of previous studies [15, 21, 58, 62, 63], it can be concluded that the populations of tropical bed bug in the studied areas could have a high level of resistance to pyrethroids. This situation calls for a decrease in reliance on pyrethroids and highlights the need for alternative insecticides with different modes of action or alternative approaches to make bed bug control more effective.
DNA sequencing identified the double mutation, M918I and L1014F, in all of studied populations. Because kdr and super kdr genotypes are presented together, a very high resistance is expected in the studied areas. Although the traditional bioassay in the present study showed resistance to permethrin, in order to identify the actual level of resistance, it is necessary to conduct further studies using a standard bioassay test. The occurrence of M918I and L1014F mutations signifies the emergence and expansion of strong pyrethroid resistance of tropical bed bug in different areas of Iran. Two hypotheses could explain the emergence and expansion of this pest. In the first hypothesis, the homogenous strains with super-kdr alleles, historically undergo strong PYs selection pressure, might have been introduced to Iran in the early third millennium and subsequently expanded rapidly throughout the country due to human-mediated movement. The second hypothesis for the expansion of strongly resistant strains includes the adaptive response to the selective pressure of applied PYs and their inability to control this emergent pest. Previously, various parts of the study areas were covered by OCs in the 1960s with the implementation of the malaria control program. After controlling malaria in the northern half of Iran, the southern parts were sprayed with OPs and CMs pesticides until the 1990s. A number of villages in the northern parts of the country were subjected to PYs, CMs, and OPs insecticide pressure in the 2010s, due to the implementation of parasite control plan by the Iranian Veterinary Organization, Ministry of Agriculture Jihad. Moreover, contaminated rooms were sprayed with different groups of insecticides (CMs, OPs, and PYs) by private pest control agents over the past three decades. In the studied populations, target site insensitivity and kdr-mediated resistance mechanism may be subjected to selection as DDT was introduced for the first time. Since DDT and PYs have a similar mode of action, cross resistance between them, the selection pressure has continued with the application of pyrethroids for controlling public health pests and pyrethroid-treated bed nets. However, after the control of malaria and the cessation of vector control programs, the selective pressure decreased during the last three decades. For the reason that there is not a huge selection pressure to justify the emergence of such homogeneity in the kdr and super-kdr allele, the probability of implying the adaptive response to selective pressure hypothesis is lower, and it is more likely that the homogenous strains with super-kdr allele were introduced to Iran. Although the double mutation (M918I and L1014F) kdr allele was found in populations from Australia, India, Thailand, and China [19, 25, 58], the origin of this invasive pest in Iran is under question because the corresponding gDNA sequences of the target gene are not publicly available. Additional complimentary studies with microsatellite or mitochondrial markers, such as the study of Zhao et al. [58], are suggested to clarify this hypothesis via investigating more samples from different populations. Our study was not designed to seek the point of the origin or introduction of pyrethroid resistance into bed bug populations. Nevertheless, this resistance may have a multifunctional origin, and mechanisms such as metabolic breakdown and efflux may implicate this issue [9]. Further epigenetic studies with more population and additional molecular markers are needed to come to a final conclusion on the origin of bed bug resistance in the study areas. Moreover, the presence of kdr resistance and the inability of PYs to control this emergent pest necessitate reconsidering control methodologies and applying products against this pest. Indeed, the application of the integrated pest management measures and determination of the ecological, social and biological risk factors, which play a key role in bed bug infestation, are important issues that requires to be considered in developing control programs against this invasive pest.
Multiple alignment of DNA sequence from C. hemipterus with that of C. lectularius presented the A468T and D953G mutations, beside the M918I and L1014F mutations, in studied populations. The 468 locus in the intracellular linker of the domain IS6-IIS1 is the hypervariable site in different species of insect. Therefore, the nucleotide variation in this locus may reflect the difference between two bed bug species. On the other hand, known kdr mutations in this region are rare and only E435K at the beginning and C785R mutations at the end of the I-II linker were identified in pyrethroid-resistant strains of the German cockroach, Blatella germanica [18]. Accordingly, the A468T substitution might be related to insecticide resistance in the tropical bed bug. The D953G mutation, located in the linker of the domain IIS5-S6, has been reported from pyrethroid-resistant strains of C. hemipterus in Thailand [25]. This mutation, which is equivalent to L936F mutation in corn earworm, Helicoverpa zea, has been verified to reduce the sensitivity to pyrethroids [18, 63]. The vgsc gene regions, where A468 and D953 are located, are major sites for the influence of PYs, and several point mutations clustering in these regions affect resistance to pyrethroids [9–11, 13, 15, 20, 21]. Therefore, this study highlights that the D953G and A468T substitutions could have additive effects on main kdr mutations in pyrethroid resistance, though other forms of resistance (cuticular and detoxification) cannot be excluded. Ultimately, the function of the A468T and D953G variations needs to be tested by the construction of mutant transcript in further studies, to confirm that this construction is actually a kdr mutation against the pyrethroid resistance.
Unlike the pyrethroid-resistant strains of C. lectularius in the present study, the L925I, I936F, V419L, and other putative kdr mutations in the IS6 and the partial domain I-II linker were not found in C. hemipterus. The existence of M918I and L1014F mutations in the tropical bed bug expresses the specific responses of this species to the selective pressure of PYs. These adaptive responses may be related to the innate immune system and DNA damage repair pathways in C. hemipterus [64, 65].
The aim of this investigation was to determine the kdr mutation in different populations of tropical bed bug, and, therefore, several point mutations, including A468T, D953G, M918I, and L1014F, were identified. Similar to the finding of our study, previous investigations have documented the D953G, M918I, and L1014F mutations in other populations of bed bugs and other insects [9, 13, 15, 25, 60, 61]. However, owing to the mechanisms of post-transcriptional regulatory variation in organisms, mRNA editing is possible [66]. Perhaps, the RNA sequence can be altered from the encoding DNA template, which can potentially lead to a change in the actual DNA expression of the kdr mutation. There are known surveys showing that insects use RNA editing to create variations in the vgsc gene [67, 68]. Consequently, future studies based on both DNA and cDNA sequencing would strengthen the findings of this study.
The results of the current study showed the high frequency of kdr mutations and pyrethroid resistance in different populations of tropical bed bug in the studied areas. Developing resistance management plans and applying effective tactics [2, 5, 69] are essential to lessen the bed bug infestation and frequency of mutant alleles. Accordingly, the use of insecticides with different mode of actions and intensive application of pyrethroids with insect growth regulators (IGRs) are recommended to control the tropical bed bug. Moreover, integration of non-chemical methods [70], i.e. applying multidisciplinary control methods to prevent pest entry, limiting access to fed on hosts and shelters, vacuuming, using impregnated covers and mattresses, and employing heat in form of dry steam and cold, are suggested for the management of this invasive pest.
Considering the fact that the passive host/furniture-mediated dispersal is the common modus operandi, and the active dispersal is restricted to within buildings, beside the mentioned control tactics, there are some other strategies that have to be viewed in developing control plans, such as educating travelers to recognize bed bugs and avoid carrying them from place to place in infested luggage, training residents to identify pest and take necessary actions when infestation is suspended, providing guidelines for accommodation suppliers in how to make rooms less bed bug friendly, and establishing instructions and rules for importing and trading of second-hand goods, furniture, and clothing [2, 63, 71]. Altogether, in setting up management plans for bed bugs, the biological and socioeconomic risk factors, as well as the side effects of current control measures against public health pests, should be taken into account.
{kind=link}