CMA Improves Cognition and Blood Parameters in Alzheimer’s Disease Patients in a randomized double-blinded clinical trial
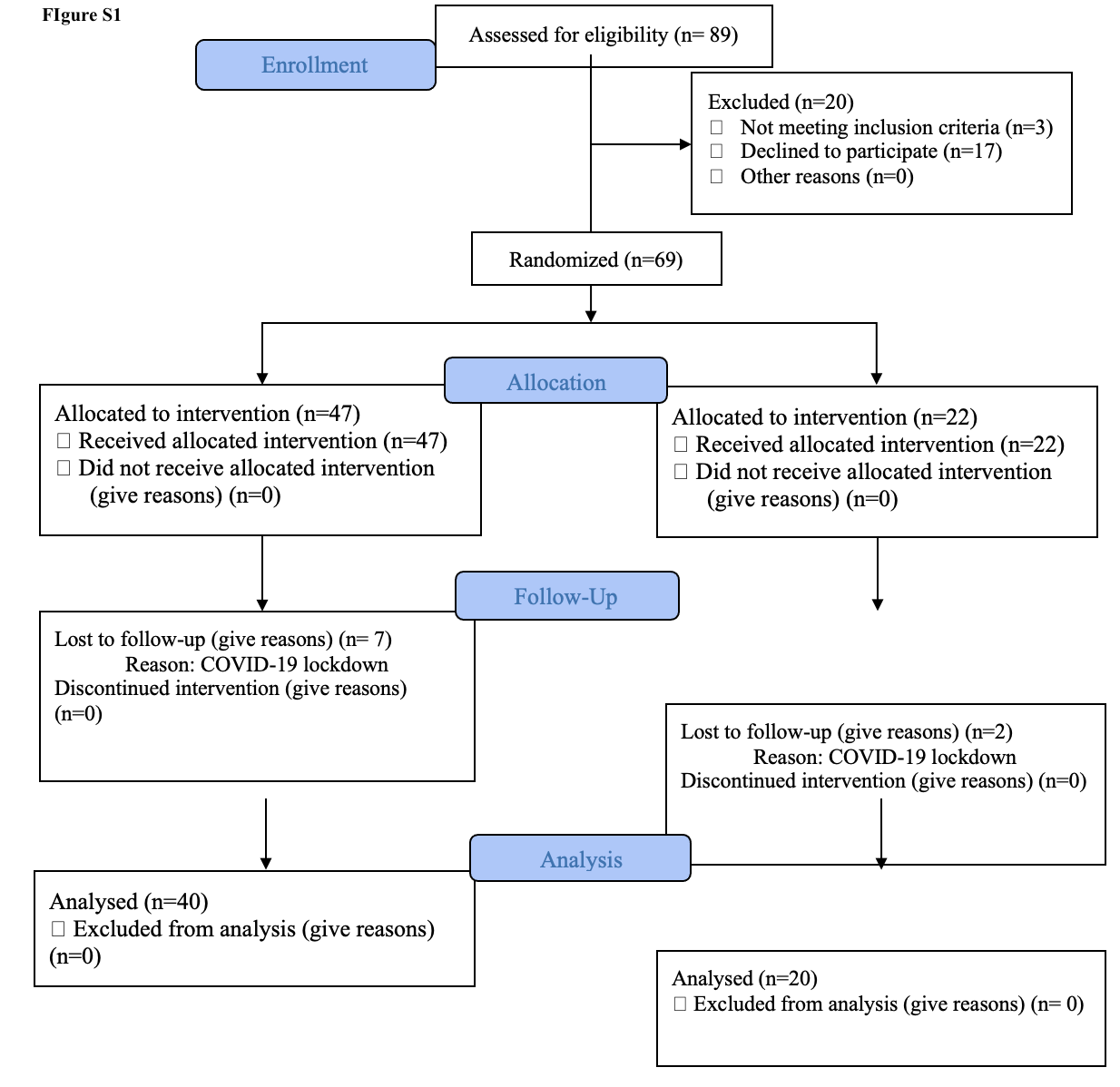
To test the effect of the CMA in AD patients, we performed a double-blind, randomised, placebo-controlled phase 2 study and screened 89 adults diagnosed with AD. We recruited 69 patients older than 50 years with mild to moderate AD according to ADAS-Cog (AD Assessment Scale-cognitive subscale; ADAS ≥ 12) and the Clinical Dementia Rating Scale Sum of Boxes (CDR-SOB; CDR ≤ 2) between 01.02.2020–01.10.2020. Of the 69 patients, 47 were randomly assigned to the CMA group and 22 to the placebo group and completed visit 2 after 28 days. Nine patients (7 in CMA, 2 in placebo groups) dropped out of the study before Day 84 visit during the COVID-19 lockdown. Of these patients, 60 (40 in the CMA group and 20 in the placebo group) completed visit 3 after 84 days (Fig. 1A, Figure S1). We assessed the clinical variables on Days 0, 28 and 84, and analysed the differences between the CMA and placebo groups (Dataset S1 and S2).
The patients' mean age in the study was 70.8 years (56–86 years), and 52.1% were men (Table 1, Dataset S1). The mean ADAS-Cog score was 22.88 (± 10.51) for CMA and 26.28 (± 17.35) for placebo (Table 1, Dataset S2). There was no significant difference for the baseline levels of these demographic parameters and AD clinical indicators between CMA and placebo groups (Table 1). The other clinical characteristics were similar in the CMA and placebo groups (Dataset S1 and S2). Regarding safety, no severe adverse events occurred, and 5 patients (7.2%) reported adverse events. All decided to complete the study (Table 2).
Table 1
Demographics and baseline characteristics of the study population*
| |
CMA (n = 47)
|
Placebo (n = 22)
|
P value
|
|
Age
|
70.77 ± 8
|
70.91 ± 7.54
|
0.97
|
|
Gender
|
|
|
|
|
Male
|
25 (53.2%)
|
11 (50%)
|
0.88
|
|
Female
|
22 (46.8%)
|
11 (50%)
|
|
Ethnicity
|
Caucasian (100%)
|
Caucasian (100%)
|
-
|
|
Body Mass Index
|
29.04 ± 5.25
|
27.43 ± 5.05
|
0.23
|
|
ADCS-ADL
|
55.79 ± 15.02
|
56.67 ± 15.63
|
0.83
|
|
ADAS-Cog
|
22.88 ± 10.51
|
26.28 ± 17.35
|
0.32
|
|
MMSE
|
19.45 ± 4.21
|
17.95 ± 5.66
|
0.22
|
|
CDR
|
0.82 ± 0.45
|
1.02 ± 0.6
|
0.09
|
|
* Presented as Mean ± Standard deviation, except gender and ethnicity.
MMSE: Mini Mental State Examination; ADAS-cog: Alzheimer's Disease Assessment Scale-cognitive subscale; ADCS-AD: Alzheimer's Disease Cooperative Study - Activities of Daily Living; CDR: Clinical Dementia Rating Scale
|
Table 2
|
Patient No
|
Treatment
|
Adverse Event
|
SOC
|
AE Intensity
|
Relationship to IMP
|
|
TR10006
|
Active
|
Pruritus
|
Skin and subcutaneous tissue disorders
|
Mild
|
Unrelated
|
|
TR10011
|
Active
|
Dizziness
|
Nervous system disorders
|
Mild
|
Unrelated
|
|
TR10034
|
Active
|
Diarrhea
|
Gastrointestinal disorders
|
Moderate
|
Unrelated
|
|
TR10036
|
Active
|
Diarrhea
|
Gastrointestinal disorders
|
Moderate
|
Unrelated
|
|
TR20045
|
Active
|
Nausea
|
Gastrointestinal disorders
|
Moderate
|
Unknown
|
We measured clinical variables in all patients and analysed the differences before and after administration in the active and placebo groups (Fig. 1B, Table 3, Dataset S2). As decreased ADAS-Cog score is the indicator of the improved cognitive function in AD patients, the ADAS-Cog scores is significantly decreased on Day 28 vs Day 0 (Log2FoldChange (FC)= -0.33, (26% improvement), p-value = 0.0000003, effect size= -0.43, 95% CI= [-0.59, -0.29]) and further decreased on Day 84 vs Day 0 (Log2FC= -0.37, (29% improvement), p-value = 0.00001, effect size= -0.50, 95% CI= [-0.71, -0.29]) in the CMA group. A slightly but significant improvement is also found in the placebo group on Day 28 vs Day 0 (Log2FC= -0.16, (12% improvement), p-value = 0.009, effect size= -0.16, 95% CI= [-0.27, -0.05]) and Day 84 vs Day 0 (Log2FC= -0.19, (14% improvement), p-value = 0.001, effect size= -0.23, 95% CI= [-0.35, -0.11]). There was no significance between groups on Day 28 and Day 84. This could be related to the placebo effect which is apparent in the early stages of AD clinical trials showing relatively unchanged response or even improvement that may continue for up to 12 months, followed by natural disease progression overtime when the drug effect becomes more prominent and the placebo response decreases (40).
Table 3 Differences in ADAS-Cog, ADCS-ADL and MMSE scores in the CMA and placebo groups

MMSE: Mini Mental State Examination; ADAS-cog: Alzheimer's Disease Assessment Scale-cognitive subscale; ADCS-AD: Alzheimer's Disease Cooperative Study - Activities of Daily Living; ADAS-Cog score > 20 is high, ≤20 is low; MRI: Magnetic resonance imaging
We also analysed the differences between clinical parameters by stratifying the patients into low-scored and high-scored ADAS-Cog groups (> 20 ADAS-Cog score is high, n = 30; ≤20 score is low, n = 39). There was a significance (Log2FC= -0.77, p-value = 0.0073, effect size= -1.23, 95% CI= [-0.3,-2.17]) between CMA and placebo groups in patients with higher ADAS-Cog scores on Day 84 (Fig. 1B, Dataset S2). Moreover, we found a significant improvement of ADAS-Cog scores between Day 28 vs Day 0 (Log2FC= -0.31, (24% improvement), p-value = 0.002, effect size= -0.48, 95% CI= [-0.76, -0.2]) and Day 84 vs Day 0 (Log2FC=-0.38, (30% improvement), p-value = 0.003, effect size= -0.59, 95% CI= [-0.97, -0.21]) in the severe CMA group and no significance difference in the severe placebo (p > 0.05 in both time points) group (Fig. 1B, Dataset S2). As shown in Fig. 1B, we observed a significant difference in the baseline value distribution and mean of ADAS-Cog scores in the severe (ADAS-Cog > 20) CMA and placebo groups due to the randomisation of the subjects. To verify our results, we selected 10 patients from the CMA group with matched ADAS-Cog values to the placebo group (p-value: 0.693) and presented the ADAS-Cog scores in Fig. 1C. We recalculated the differences in ADAS-Cog scores and again found significant improvement in the CMA group, whereas we found no significant difference in the placebo group. Our results indicated that the severe AD patients with high ADAS-Cog scores are more responsive to CMA.
Other primary endpoints were Alzheimer's Disease Cooperative Study - Activities of Daily Living (ADCS-ADL) and Mini Mental State Examination (MMSE). No significance was found between time points and between groups in ADCS-ADL (Fig. 1B, Table 3, Dataset S2). For MMSE, a significance in the placebo was seen when comparing Day28 and Day84 to Day0 (p-value = 0.04 and p-value = 0.02, respectively) as well as in CMA group when comparing Day 28 to Day 0 (p-value = 0.02) but not in Day84 to Day0 (Fig. 1B, Table 3, Dataset S2). There was no significance in ADCS-ADL and MMSE in any time points and between groups in patients evaluated with MRI (Dataset S2).
Analysis of secondary outcome variables showed that serum alanine aminotransferase (ALT) levels (Log2FC=-0.38, p-value = 0.01) and the uric acid levels (Log2FC=-0.19, p-value = 0.001) were significantly lower on Day 84 vs Day 0 only in the CMA group (Fig. 1D, Dataset S2). This reduction was seen both in high- and low-ALT level groups. In contrast, we found no significantly altered parameters on Day 84 vs Day 0 in the placebo group (Fig. 1D, Dataset S2).
We also measured the complete blood count parameters and found that their levels were significantly changed in the CMA group (Fig. 1D, Dataset S2). We found that the levels of platelets, basophil % and absolute numbers of basophil and neutrophil were significantly lower on Day 84 vs Day 0 only in the CMA group. In contrast, we found that the levels of monocytes were significantly increased on Day 84 vs Day 0 in the CMA group (Fig. 1D, Dataset S2). Hence, our analysis indicated that the administration of CMA improved the clinical parameters in parallel to the improvement in cognitive functions in AD patients.
Blood profile informs the response to CMA
Treatment response variability and clinical heterogeneity in AD are well documented in the literature. We observed interindividual variability in clinical measures in responses to CMA administration. Therefore, we hypothesised that some patients would respond better to CMA than others and that clinical measurements could define these subsets.
To determine whether alanine transferase (ALT), a marker for liver damage, could indicate a better response to CMA, we stratified the patients into high and low ALT groups by the median ALT of all patients on Day 0. As shown in Fig. 2A, the patients of the CMA group with low ALT achieved a significant improvement in ADAS-Cog score over different time points, while the patients in the placebo group had no improvement. In contrast, the patients of the CMA group with high ALT levels also exhibited an improved (i.e., decreased) ADAS-Cog score, but the degree of change was not as much as the patients in the CMA group with low ALT levels. Moreover, patients in the placebo group with high ALT levels also had improved ADAS-Cog scores. Thus, these results suggest that the patients with low ALT levels are more responsive to CMA.
We repeated this stratification for each blood parameter to determine the patient conditions in which CMA produces a tremendous response (Fig. 2B). In addition to low ALT, we identified high alkaline phosphatase (ALP), low gamma-glutamyl transferase (GGT), high hematocrit, high HbA1c, high insulin, high uric acid, high basophil count, and high red blood cell count as indicators for better responsiveness to CMA.
CMA Increases the Plasma Levels of Metabolites Associated with Metabolic Activators
We first analysed the plasma levels of serine, carnitine, NR, cysteine and their by-products. CMA administration increased the plasma levels of metabolic activators on Day 84 vs Day 0 in the CMA group (Fig. 3A, Dataset S3). Moreover, the plasma levels of NR, 1-methylnicotinamide, nicotinurate, N1-methyl-2-pyridone-5-carboxamide and nicotinamide (associated with NR and NAD + metabolism); of serine, glycine and sarcosine (associated with serine and glycine metabolism); and of deoxycarnitine and carnitine (associated with carnitine metabolism) were significantly higher in the CMA group on Day 84 compared to Day 0.
Next, we investigated the relationship between the plasma level of administrated metabolic activators and other metabolites. We analysed 195 of the most significantly correlated plasma metabolites with serine, L-carnitine, NR, and cysteine (Dataset S4). We found two clusters of metabolites that are significantly correlated with cysteine only or together with serine, carnitine and NR. We observed that cysteine had different plasma changes than the other three metabolic activators, as reported in previous clinical trials (29, 41).
Effect of CMA on Global Metabolism
We identified the significantly (FDR < 0.05) different plasma metabolites on Day 84 vs Day 0 and found that the plasma levels of 132 metabolites were significantly different in the CMA group (Fig. 3, Dataset S3). Evaluation of plasma metabolites that differed significantly on Day 84 vs Day 0 in each group showed that a larger number of metabolites related to amino acid (n = 53), lipid metabolism (n = 42) and other metabolic pathways (n = 37) were altered in the CMA group compared to the placebo group (Fig. 3, Dataset S3).
N-acetyl aspartate (NAA) is one of the most abundant brain metabolites, and its reduced plasma levels are associated with brain tissue damage. Previous research revealed the importance of NAA in maintaining energy metabolism in the central nervous system (42). We observed that plasma levels of NAA significantly increased on Day 84 vs Day 0 in the CMA group (Fig. 3B, Dataset S3). Another upregulated metabolite on Day 84 vs Day 0 in the CMA group is sarcosine (a derivative of glycine) which has been widely studied for its improving effects on cognitive symptoms by different pharmacological activities in neurons (43). Of note, quinolinic acid (an endogenous excitotoxin acting on N-Methyl-D-aspartate receptors leading to neurotoxic damage) levels significantly decreased on Day 84 vs Day 0 only in the CMA group (Fig. 3B, Dataset S3).
Increased plasma homocysteine levels are a known risk factor for AD, and several animal studies implicated the promising results of methionine restriction (44, 45). In our clinical trial, plasma levels of S-adenosylhomocysteine, 2,3-dihydroxy-5-methylthio-4-pentenoate (DMTPA) and N-acetyl taurine were significantly downregulated on Day 84 vs Day 0 in the CMA group (Fig. 3B, Dataset S3). Of note, reductions of these metabolites are significantly correlated with serine and NR supplementation (Fig. 4A, Dataset S4).
Increased plasma levels of metabolites in the kynurenine pathway are associated with AD severity (44). In our study, we found that plasma levels of kynurenate and 8-methoxykynurenate were significantly lower on Day 84 vs Day 0 in the CMA group (Fig. 3B, Dataset S3). Reduction in the plasma level of kynurenate was positively correlated with plasma serine levels (Dataset S4). Kynurenate, which has a prooxidant effect, is the product of the tryptophan degradation pathway. Its aerobic irradiation produces superoxide radicals and leads to cytochrome C reduction (46). It has been reported that increased levels of kynurenine lead to cell death through the reactive oxygen species (ROS) pathway in nature killer (NK) cells (47) and lower blood pressure in systemic inflammation (48).
Emerging evidence indicates a link between abnormal kidney function and AD, but the potential impact of the kidney on cognitive impairment is still undetermined (49). Recent studies showed that plasma levels of N,N,N-trimethyl-5-aminovalerate are involved in lysine metabolism, and it serves as an indicator of elevated urinary albumin excretion (50). Here, we found that the plasma level of N,N,N-trimethyl-5-aminovalerate was significantly decreased on Day 84 vs Day 0 in the CMA group (Fig. 3B, Dataset S3) and significantly inversely correlated with the plasma level of serine and NR. Moreover, the plasma level of creatinine was also significantly decreased on Day 84 vs Day 0 in the CMA group (Fig. 3B, Dataset S3). The plasma reduction in creatinine is inversely correlated with the plasma level of serine (Dataset S4). Additionally, our analysis revealed decreased levels of several metabolites belonging to histidine metabolism in the CMA group on Day 84 vs Day 0. Among those N-acetyl-1-methylhistidine is associated with chronic kidney disease and showed a significant negative correlation with serine supplementation (Fig. 3B, Dataset S4). Also, we found that plasma levels of metabolites related to the urea cycle (3-amino-2-piperidone, arginine, homoarginine, N-alpha-acetylornithine, ornithine and pro-hydroxy-pro) were significantly decreased in the CMA group on Day 84 vs Day 0 (Fig. 3B, Dataset S3) and inversely correlated with the plasma level of serine and NR (Dataset S4).
Lipids play a fundamental role in the pathophysiology of neurodegenerative diseases, including AD. Specific lipid species of cellular membranes (e.g., cholesterol and sphingolipids) are structural components of cell membranes and regulate many critical aspects of brain functions(51). In our study, plasma levels of many metabolites associated with sphingomyelins and fatty acid metabolism (acyl carnitines) were significantly increased on Day 84 vs Day 0 in the CMA group (Fig. 3C, Dataset S3). Interestingly, plasma levels of pregnenolone steroids and 2R,3R-dihydroxybutyrate were significantly decreased on Day 84 vs Day 0 (Fig. 3C, Dataset S3). These alterations were significantly positively correlated with carnitine and serine levels (Dataset S4).
Effect of CMA on Plasma Proteins
Plasma levels of 1466 protein markers were measured with the plasma proteome profiling platform Proximity Extension Assay quantifying the plasma level of target proteins. After quality control and exclusion of proteins with missing values in more than 50% of samples, 1463 proteins were analysed (Dataset S5). Proteins whose levels differed significantly between the visits in the CMA and placebo groups are listed in Dataset S5.
We analysed the effect of CMA on plasma protein profile and found that 22 proteins were significantly (p-value < 0.01) different in the CMA group on Day 84 vs Day 0. Nineteen of these proteins were significantly decreased, whereas 3 of these proteins were significantly increased on Day 84 vs Day 0. After filtering out the proteins based on log2FC, we found that the plasma levels of PSPN, OSM, PADI4, PDGFC, SCGN, LTBP3, CLEC4G, MERTK, WNT9A, ISM1, ASAH2, CES3, HPGDS, NPY, THPO, SIGLEC6, GDNF, PADI2 and EGFL7 were significantly downregulated in the CMA group. The plasma level of KLB, BGN, and ST3GAL1 was significantly upregulated in the CMA group (Fig. 4B, Dataset S5). We observed that only one significantly (p-value < 0.01) altered protein - EGFL7 upregulated- in the placebo group (Fig. 4B, Dataset S5).
The proteomic analysis in this study revealed significant alteration in levels of several critical proteins that play an essential role in the pathogenesis of AD. For instance, levels of MertK (52, 53), EGFR (54, 55), oncostatin (56–60), PAD4 (61, 62), LTGF (63–67), and TPO (68), known as a potent inducer of neuro-inflammation, amyloid production and apoptosis, decreased. In contrast, proteins with neuroprotective and pro-cognitive properties, such as Klotho (69, 70) and ST3GAL1 (71), increased after CMA treatment. More interestingly, most of the analysed proteins were also significantly altered in recent human AD studies (63–67, 72–78). KlothoB levels were also significantly altered after CMA treatment, consistent with their neuroprotective role as a cofactor and neurotrophic factor. Recent studies have shown that KlothoB indirectly regulates glucose and energy metabolism through F2F1, expressed in some regions of the brain involved in learning and memory (77). Moreover, GABA signalling has also been shown to play a critical role in mediating the detrimental effects of increased dihydroxybutyrate levels in the progression of MCI (79). Interestingly, our metabolomic study indicated decreased post-therapeutic dihydroxybutyrate levels. Although the exact pathways involved in the metabolic generation of DHBA are still far from clear, it has been hypothesised that dihydroxybutyrate levels may be a compensatory response to increased cellular stress secondary to compromise of the Krebs cycle function, creating an alternative energy production pathway in AD (79). This represents indirect evidence to suggest that our treatment exhibits an energetic regulatory function.
Integrative Multi-Omics Analysis
Multi-omics data integrations have been proven to give novel insights and a more holistic view of the human body in both health and disease states (80). In this study, we generated an integrative multi-omics network using metabolomics and proteomics data, coupled with detailed clinical variables, to understand the functional relationships between analytes from the same and different omics data types. We generated the network using the method used in iNetModels (39), to which we also deposited our network. The network consists of 937,282 edges from 2,273 nodes (36.3% network density, Dataset S6).
We extracted a sub-network to highlight the interactions between the individual metabolic activators, cognitive function (ADAS-Cog scores), two highlighted proteins (OSM and PSPN), and their top neighbours (Fig. 4C). From the sub-network, ADAS-Cog was negatively associated with carnitine (and its derivatives) and nicotinamide associated metabolites, whereas the metabolic activators were negatively associated with fatty acid and histidine metabolism. Finally, we observed that, among others, OSM and PSPN were positively associated with immune and cell cycle-related proteins.
Subsequently, we performed centrality analysis to identify the most central analytes in the networks. The top 20 most central metabolites were dominated by amino acid metabolites (tryptophan, glutamate, and branched-chain amino acid metabolism) and lipid metabolites (androgenic steroid pathway), where top proteins were related to, among others, short- and long-term memory (CALB1), lipid metabolism (PLA2G10), and immune response (SELPLG, CLEC4D, and LGALS7).
Furthermore, we performed community analysis within the network using the Leiden algorithm. We discovered 3 modules that showed significant interaction among the members. In cluster-0, the biggest clusters, the top nodes were related to tryptophan metabolism (indole acetate), fatty acid metabolism (3-hydroxyoctanoate), and steroid metabolism (11-ketoetiocholanolone glucuronide and 11-beta-hydroxyetiocholanolone glucuronide). Moreover, we found 2 top proteins in the same cluster, ACTA2 and IGFBP1, associated with AD (81, 82). In cluster-1, the top nodes were associated with leucine metabolism (3-hydroxy-2-ethylpropionate), ceramide phosphatidylethanolamine, and a carnitine metabolite (erucoylcarnitine); meanwhile, cluster-2’s central nodes were related to methionine metabolism and aminosugar metabolism (N-acetylglucosamine/N-acetylgalactosamine). These results showed that the integrative multi-omics network analysis could be used to strengthen the results from single omics analyses and identify key analytes associated with AD. Moreover, it provided new insights by elucidating the functional relationships within and between different omics data.
In evaluating the correlations between each activators (used in the present study for therapeutic purposes) and clinical, metabolic, and proteomic parameters, we identified significant correlations between serine, carnitine, cysteine, and nicotinamide levels and improved peripheral blood parameters, such as liver function, CBC, and glycated hemoglobin (HbA1c), which are relevant to the pathogenesis of AD. Accordingly, improved ADAS-Cog scores were also associated with serum serine and carnitine changes, which fit well with their well-known pro-cognitive and energy-boosting effects. Similar results were also observed for metabolomic and proteomic data. The majority of the activators exhibited significant correlations with improved metabolites and proteins (either increased or decreased) relative to a slower degeneration process in AD. It is worth mentioning here that two of the proteins, OSM and PSPN, most strongly associated with other beneficial protein metabolites, were also related to several critical amino acid alterations, such as spermidine and hypotaurine, which may suggest a metabolic shift from the protein to amino acid metabolism to compensate the energy deficit reported in AD.
Effect of CMA on Hippocampal volumes and Cortical Thickness
The MRI group's baseline demographics and improved clinical parameters significantly aligned with the entire patient cohort (Dataset S1&S2). Based on comparing the differences between pre and post-treatment groups in each treatment arm (83), our results showed that the left whole hippocampal mean volume, and left molecular layer HP body were significantly improved after the CMA treatment (p < 0.05, Fig. 5B, Dataset S7). Several other sub-anatomic hippocampal regions (left CA1 body, left whole hippocampal body) were improved near to a statistically significant level (p < 0.05, Fig. 5B, Dataset S7) after the CMA treatment. According to longitudinal cortical thickness analysis, the active group showed statistically significant alterations in the bilaterally lateral occipital, bilateral rostral middle frontal, left inferior parietal, and left paracentral regions (Fig. 5C).
{kind=link}