Furin and related PCs (PC2, PC1/3, PC4, PACE4, PC5/6, and PC7) are specialized serine endoproteases that cleave the multibasic motifs R-X-(R/K/X)-R↓(S)((V/A/L) 9, 10 11. In addition to its normal cellular functions, furin is also implicated in many pathogenic states. Thus, furin cleaves to maturity membrane fusion proteins of viruses and pro-toxins of a variety of bacteria, including anthrax and botulinum toxins, influenza, measles, flaviviruses and many others 9, 20. Acquisition of furin-like priming sequences correlates with increased virulence and pathogenicity. For example, the acquisition of a furin-cleavage site in the priming site of the viral protein hemagglutinin (HA), necessary for influenza virus entry, is associated with the increased pathogenicity of the avian influenza virus 21. Perhaps more interestingly, such evolution of the influenza virus to contain furin–like sequences can be induced by repeated passages in cell culture or through animals 22.
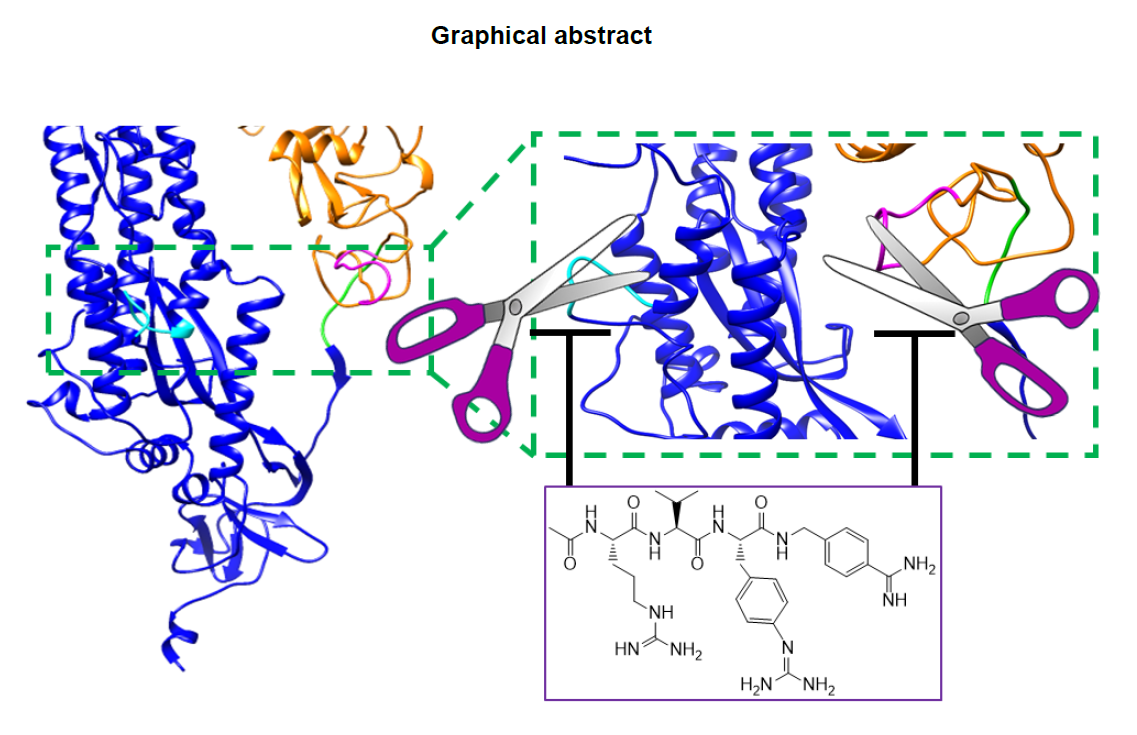
Furin-like sequences that may contribute to increased virulence have also been identified in the coronavirus spike glycoproteins (Table 1)1, 2, 3, 8, 12, 23. The complex mechanism of viral fusion in coronaviruses is not fully elucidated, but it likely comprises a first cleavage of the S1 site that allows the S2 subunit to more easily dissociate from the S1 subunit (Figure 1).. The S2 subunit contains a fusion peptide, an internal fusion peptide, two heptad-repeat domains, and a transmembrane domain (Figure 1).. The spike protein S1 attaches the virion to the cell membrane by interacting with its host receptor, thus initiating the infection. This occurs most likely by binding to the ACE2 receptor causing internalization of the virus into the endosomes of the host cell. Proteolysis by serine-proteases of the S1 site (or by cathepsin L, in the adjacent S2 cleavage site)8 may unmask the fusion peptide and activate membrane fusion within the endosomes. This step seems to require an additional cleavage at the S2’ site to unmask the internal fusion peptide in the S2 viral fusion protein.
However, while the S2 site is conserved among various coronavirus strains, the S1 site in SARS-CoV2 contains a furin cleavage site (Table 1).. The S1 sequence is located in an exposed unstructured loop in the structure of the SARS-CoV2 spike protein (Figure 1A,B).. As a result, no electron density was observed in this loop region in the recently reported Cryo-EM structure1. Most intriguingly, the unusual SARS-CoV2 S1 site may have also acquired an increased cleavage propensity for TMPRSS2 (Table 1) (supplementary material). This may explain why TMPRSS2 appeared more important than furin for the entry of surrogate SARS-CoV2 viral particles in cell 4. However, while TMPRSS2 is abundant in the respiratory tract, furin is more ubiquitous in many other organs, (https://www.proteinatlas.org/ENSG00000140564-FURIN/tissue),, hence acquisition of a furin cleavage site most likely increases the tropism and the pathogenicity of the strain. Furthermore, because furin is localized in the trans-Golgi network and cycles between the trans-Golgi and the cell surface, furin cleavage in the spike protein may occur also during viral egress from the infected cells. As a result, the pre-primed virus may be more ready to enter and infect other cell types and to spread among hosts.
The common mechanisms of cell trafficking mediated by furin cleavage by both viral fusion proteins and bacterial toxins is striking. For example, anthrax toxin, similar to SARS-CoV2, requires processing of the PA sequence RKKRST (Table 1),, to chaperone the internalization of the LF toxin into macrophages. Intriguingly, much like the S1 of SARS-CoV2, the PA cleavage site also contains both furin-like and TMPSS-like proteases recognition sites (Table 1),, and it also invades macrophages, making a potentially good model system to study inhibition of priming in vivo. Intact toxins, like viral proteins, are incapable of accomplishing these processes in absence of proper priming by the host proteases. In recent years, several reports emerged describing improved furin inhibitors24, 25, 26, 27, 28. Here, we evaluated the potential of systemic administration of a furin protease inhibitor to prevent priming, using the serine protease dependent anthrax toxin as a model system.
When tested in cell, the pan-active compound 1 was efficacious in protecting RAW macrophages from anthrax toxin (Figure 2),, suggesting that the prototype agent possesses favorable pharmacological properties for in vivo studies (supplementary material). Because the toxicity in vivo of LF/PA toxin intimately depends on PA cleavage by serine proteases,19 this model is ideal for evaluating the inhibition of priming in vivo. Hence, Balb/C mice receiving a mixture of LF and PA (100 μg via the tail vein) were injected with either a single dose 3 mg/kg (I. P.) of compound 1, or two doses spaced by 2 h, or by vehicle control. According to our pharmacokinetics studies, these doses should reach blood levels of the drug sufficient to inhibit furin effectively (Figure 2B).. Based on previous studies19, mice treated with such a lethal dose of toxin die by roughly 48 h post-treatment depending on the LF and PA lots, the age and strain of mice and their weight. A potent direct LF inhibitor given at 30 mg/kg I. P. 3 times a day was reported to rescue death of mice at 48 h, while no survivors were present in the control group29. However, no information was provided in the literature on the fate of treated mice after 48 h (no time to death was reported). In our experiments, all mice in the untreated group died by time = 33 hr, in close agreement with the published studies, while a remarkable and significant increase of both median survival time (MST) and time to death (TTD) was observed in both groups treated with compound 1 (Figure 3B) even at the single dose of 3 mg/kg.
Recently, camostat mesylate, a covalent TMPRSS2 inhibitor already clinically approved for other indications in Japan (Figure 2D),, has been proposed given that it partially blocked viral entry in surrogate cellular assays 4. However, we found that camostat mesylate did not appreciably inhibit furin (Figure 2E),, and while it may attenuate entry at relatively high concentrations (10- 50 μM)4, in our opinion it would do little to prevent furin-mediated egress of partially primed (at S1 site), hence more virulent, SARS-CoV2 viral particles.
Hence, while we await for the results of the efficacy of camostat in a very recently initiated human clinical trial with COVID19 patients (https://clinicaltrials.gov/ct2/show/NCT04321096), this report wishes to incentivize once again private and public efforts to consider developing additional pan-serine protease inhibitors that target also furin, perhaps taking advantage of several agents that have already been reported in pre-clinical studies24, 25, 26, 27, 28, into emergency therapeutics to combat the new coronavirus SARS-CoV2, and to ward off future similar pandemics that are likely to occur when pathogens acquire the further optimized furin cleavage sites within their priming entry mechanisms. These development efforts are particularly significant especially for coronaviruses as no viable treatments or vaccines are currently available, and at the same time other future furin-like cross-species transmission in coronaviruses seems likely. Mutations of the cleavage site in either S1 or S2’ of coronavirus strains’ spike glycoprotein can be correlated with pathogenicity, increased tropism, and crossing zoonotic barriers. Unfortunately, one could envision several mutations in the SARS-CoV2 (or any other coronavirus strain) that could transform these sequences into more efficient furin and/or dual furin and TMPRSS2 cleavable sites, hence increasing their pathogenicity, virulence, and potential for spread. For example, the S2’ sequence for SARS-CoV2 could be more efficiently cleaved by furin if a single nucleotide mutation would occur to substitute Pro for an Arg residue (Pro/Arg; Pro is coded by GGA, GGG, GGT, GGC while Arg is coded by GCA, GCG, GCT, GCC); that is, if the current S2’ sequence SKPSKRSF (furin cleavage score 0.42) would turn into the sequence SKRSKRSF (furin cleavage score 0.81) (Table 1, supplementary material). Further mutations could also be easily envisioned that would introduce a second TMPRSS2 cleavage site. For example, a S2’ sequence SKRKKR↓SF would contain both a furin site (furin cleavage score 0.86) and a preferred R↓K site for TMPRSS2-like cleavage (Table 1),, just to cite few possibilities. Luckily, analysis of currently deposited sequences of SARS-CoV2 (https://www.ncbi.nlm.nih.gov/genbank/sars-cov–2-seqs) revealed that these mutations have not occurred to date.
{kind=link}