Currently, there are no truly ideal methods for optimal targeted drug treatments for PD. It has become a research hotspot for decreasing toxicity and side effects of anti-PD medications and finding alternative drugs. TCM typically showed a multi-component and multi-pathway synergistic effect in treating PD. Compound Dihuang Granule was developed associated with clinical experience in treating PD. To further study the CDG mechanism in treating PD, we screened the key active components of CDG in treating PD utilizing network pharmacology and found that the CDG mechanism in treating PD could be closely associated with the Toll-like receptor and NF-KB pathways. Therefore, the following animal experiments will verify essential proteins expression on TLR4/NF-KB signal pathway and further explore the potential molecular pathway of CDG treatment of PD.
Our previous study demonstrated that CDG played a significant neuroprotective role in the MPTP-induced PD model and the 6-OHDA-induced PD model in vivo and could effectively improve gastrointestinal dysfunction and suppress microglia activation in PD mice [17, 18, 25]. Gut microbiosis is a potential factor in PD pathogenesis, and that gut microbiota was connected to PD progression by the gut-brain axis interaction [26, 27]. Herein, intraperitoneal MPTP injection was used to establish a PD mouse model related to intestinal microbiota disorders to measure CDG neuroprotective effect on PD and further study its treatment mechanism.
PD research has thoroughly studied MPTP, among the highest-known neurotoxins disrupting dopaminergic neurons [28]. Due to its lipophilicity, MPTP might pass the blood-brain barrier, transforming it to MPDP+ by monoamine oxidase B in astrocytes. MPP+ is the active compound of dopamine neurons that enters the SNpc. It suppresses complex I of the mitochondrial electron transport chain, resulting in reduced ATP, oxidative stress, as well as degeneration of DA ergic neurons [29, 30, 31]. Herein, MPTP-induced PD mice exhibited motor dysfunctions in open-field and pole tests. Oxidative stress was crucial in PD development and might induce degeneration of DA ergic neurons. SOD, a radical scavenger, and MDA, a lipid peroxidation metabolite, are important indicators revealing oxidative stress levels [32, 33].
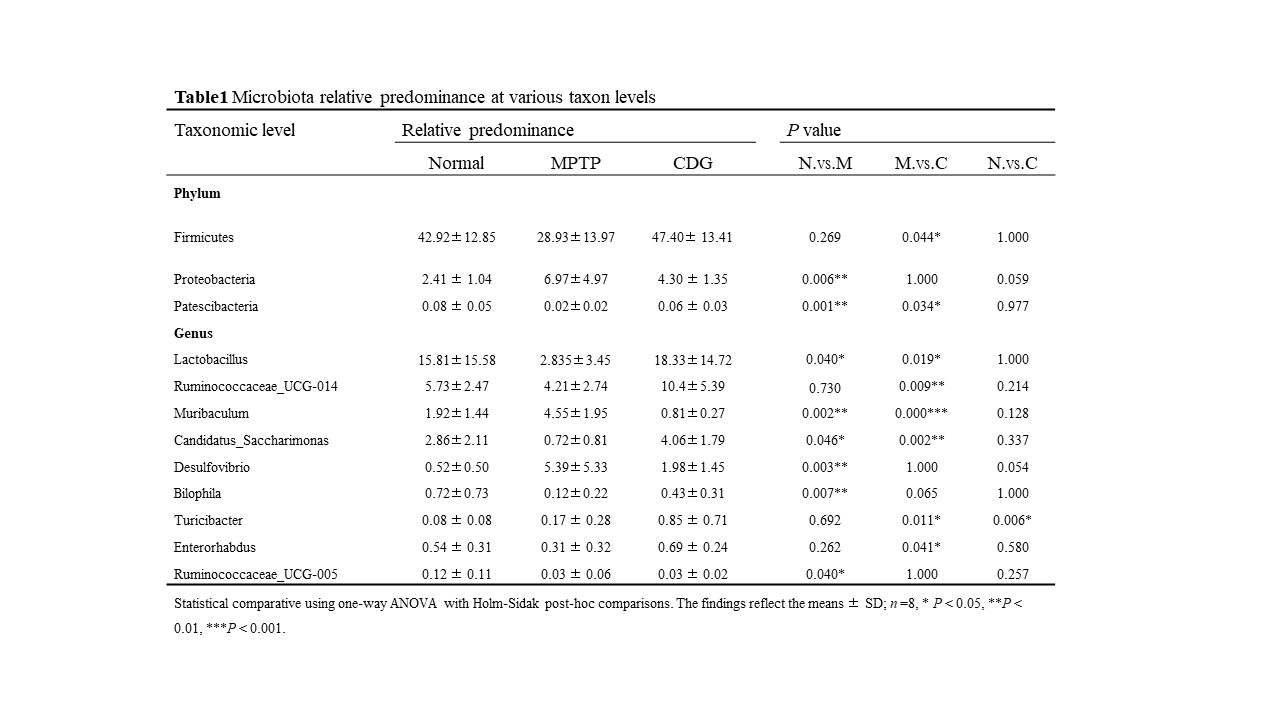
Moreover, PD mice had increased MDA and decreased SOD expression in serum, which had an oxidative stress response. IHC of brain tissue and WB showed DA damage. Together, MPTP-induced mice had PD pathology and motor symptoms. Furthermore, MPTP-induced PD mice exhibited dysbiota dysbiosis [34, 35, 36]. Therefore, we analyzed the composition of the gut microbiota in PD mice by 16s rDNA sequencing. The findings demonstrated a non-significant variation in alpha diversity among the three groups, indicating that our sample sizes should be increased. Alpha diversity showed no changes between PD patients and principal normal subjects [37]. Moreover, beta diversity changed significantly among the three groups. Proteobacteria and Patescibacteria abundance increased significantly in PD mice at the phylum level. Proteobacteria were associated with gastrointestinal inflammation, with higher content in PD patients, and increased Proteobacteria abundance in PD mice with MPTP molding [34, 38], consistent with our animal experiments. Lactobacillus is a probiotic bacterium that is reduced in PD patients and has also been reduced in MPTP-induced mice [5, 39].
Additionally, we first found that Genus Muribaculum and Candidatus_Saccharimonas were associated with the PD, with increased Muribaculum and decreased Candidatus_Saccharimonas in MPTP-induced PD mice. As per our results, the decreased Genus Bilophila abundance was associated with PD. Bilophila abundance is correlated with Hoehn and Yahr stages, revealing that Bilophila can serve as a progression indicator in PD [40]. Desulfovibrio can produce hydrogen sulfide and LPS, which may induce the aggregation of α-synaptic nuclear proteins, with a higher abundance as observed in PD patients [41], also increased at Genus herein. Briefly, MPTP-induced mice had dysbiota dysbiosis, and the mechanisms of intestinal microbiota in PD progression still need further exploration.
Further analysis of the microbiota revealed that reestablishing normal gut microflora could enable the protective effect of CDG therapy. CDG increased the abundance of Phylum Firmicutes in PD mice. Firmicutes can produce butyrate, which is involved in the inflammatory response of body [42]. SCFAs have crucial roles in promoting intestinal function and anti-inflammatory effects. Ruminococcaceae_UCG-005 and Ruminococcaceae_UCG-014 can produce SCFAs and exert their anti-inflammatory effects [43, 44]. Herein, Genus Ruminococcaceae_UCG-005 was reduced in PD mice, and Genus Ruminococcaceae_UCG-014 was increased after CDG treatment, adjusting for the microflora of PD mice. Moreover, the abundance of beneficial bacteria Enterorhabdus was increased after CDG administration. Turicibacter is associated with disordered lipid metabolism and can exert anti-inflammatory effects, and Turicibacter is positively associated with SCFA production [45]. Herein, the predominance of Turicibacter was raised in the PD mice after CDG administration. The increase of Turicibacter in the model group might be associated with the PD immunomodulation of mice. Overall, our data supported that CDG significantly altered some inflammation-associated microflora in MPTP-induced mice, increasing beneficial bacteria.
Gut microbiota and its metabolites can participate in PD pathophysiology by regulating CNS neuroinflammation, gut inflammation, and barrier function through gut-brain axis interactions [46]. Dysbiosis of gut microbes might cause bacteria to produce harmful metabolites, such as lipopolysaccharides (LPS), which could disrupt the functionality of gastrointestinal barrier. TLR4 is the primary receptor for LPS immune recognition, widespread express on the surface of several cells, including microglial, astrocyte, and intestinal epithelial cells. Its activation triggers the host inflammatory response and the proinflammatory factors, including TNF-α, IL-1β, and IL-6 [47, 48, 49]. These findings revealed that gut microbiota might drive the colonic mucosa immunological activation via the TLR4 signaling pathway, resulting in neuroinflammation and further neurodegeneration in PD [50]. To further explore the mechanism by which CDG exerts neural protection by replenishing the intestinal microbiota of PD mice, we highlighted the relationship of inflammation between the gut and the brain. Microglia and astrocytes are crucial in maintaining neuronal function and brain homeostasis and are also the primary cells mediating innate immunity in the CNS [51, 52]. When activated, they secrete large quantities of inflammatory chemokines, including TNF-α and IL-1β [53]. Astrocytes can recognize the TLR mediating inflammatory CNS disease injury [47].
Our study found that microglia and astrocytes were activated in the SNpc of PD mice and were inhibited after CDG administration. Moreover, we labeled TLR4 and TNF-α cells with IF to assess intestinal inflammation. CDG reduced the infiltration of TLR4 and TNF-α cells in PD mice colonic tissue. We also discovered that cytokines (TNF-α, IL-1β) were raised in PD mice serum, suggesting that proinflammatory factors may penetrate the systemic circulation and cause a systemic inflammatory response. Gut microbial dysbiosis-mediated inflammation leads to the intestine in high permeability to cause “intestinal leakage.” Gut barrier integrity is strongly associated with tight junction protein ZO-1 expression in intestinal tissue [24]. ZO-1 expression is lowered in PD mice, and ZO-1 expression is reduced in inflammatory bowel disease (IBD) patients [54, 55]. We examined ZO-1 expression in the mouse colon utilizing IF assay and found that CDG protected the intestinal barrier via restoring tight-junction function. All the data showed that CDG could reconstruct the intestinal microbiota of PD mice to reduce microglia and astrocytes activation, inhibit serum inflammation, and protect the damaged intestinal barrier to play a neuroprotective role.
NF-κB activating factor TANK-binding kinase 1 (TBK1) is a crucial factor of NF-κB and has a key part in the inflammatory immune reaction [49, 56]. Inflammatory cytokines stimulate NF-κB activation in TLR4 signaling through heterodimer RelA (p65) and p50, which translocate to the nucleus and interact with NF-κB target sites in immune response genes [49]. After TLR4 ligand binding, NF-κB signaling is activated to discharge proinflammatory cytokines (TNF-α and IL-1β), which has a crucial function in innate immune defense, neuronal excitotoxicity, and neurodegeneration [57]. Suppressing TLR4-mediated NF-κB signaling may be a successful approach against PD [58]. FLZ protects rotenone-induced PD mice by ameliorating intestinal dysbiosis and inhibiting TLR4 / MyD88 / NF- κB signaling pathway in SN and colon [59]. Herein, we further demonstrated the crucial function of activating the TLR4 pathway in the microbe-gut-brain axis involved in PD pathology. Besides reconstructing the dysbiota dysbiosis in PD mice, CDG effectively reduced TLR4, TBK1, NF-κB, and TNF-α expressions in the PD mice striatum and colon. The molecular mechanisms of CDG in improving dysbiota dysbiosis could be connected to inhibiting TLR4/NF-κB pathway.
{kind=link}