Engineering the Sabin 1 and 3 genomes to prevent reversion to neurovirulence
To engineer new type 1 and type 3 Sabin strains, we replaced the capsid (P1) region of the nOPV2 candidate 1 (-c1) with the capsid region from a Sabin 1 or 3 clone to generate nOPV1-c1 and nOPV3-c1 (Fig. 1A). For simplicity, the -c1 designation is omitted hereafter. This strategy preserved all the modifications introduced in nOPV2 except that the capsid proteins now corresponded to types 1 and 3. The capsid regions of all three Sabin strains contain attenuating mutations 6,32,33. Accordingly, the genome of the new polio vaccine candidates carries five modifications of the Sabin 2 genome, including two modifications within the 5′UTR (relocated cre, and S15domV), synonymous mutations at eight nucleotide positions in the 2C coding region to inactivate the internal cre34 and two mutations in the 3D polymerase (D53N, and K38R) to limit viral adaptability (Fig. 1a).
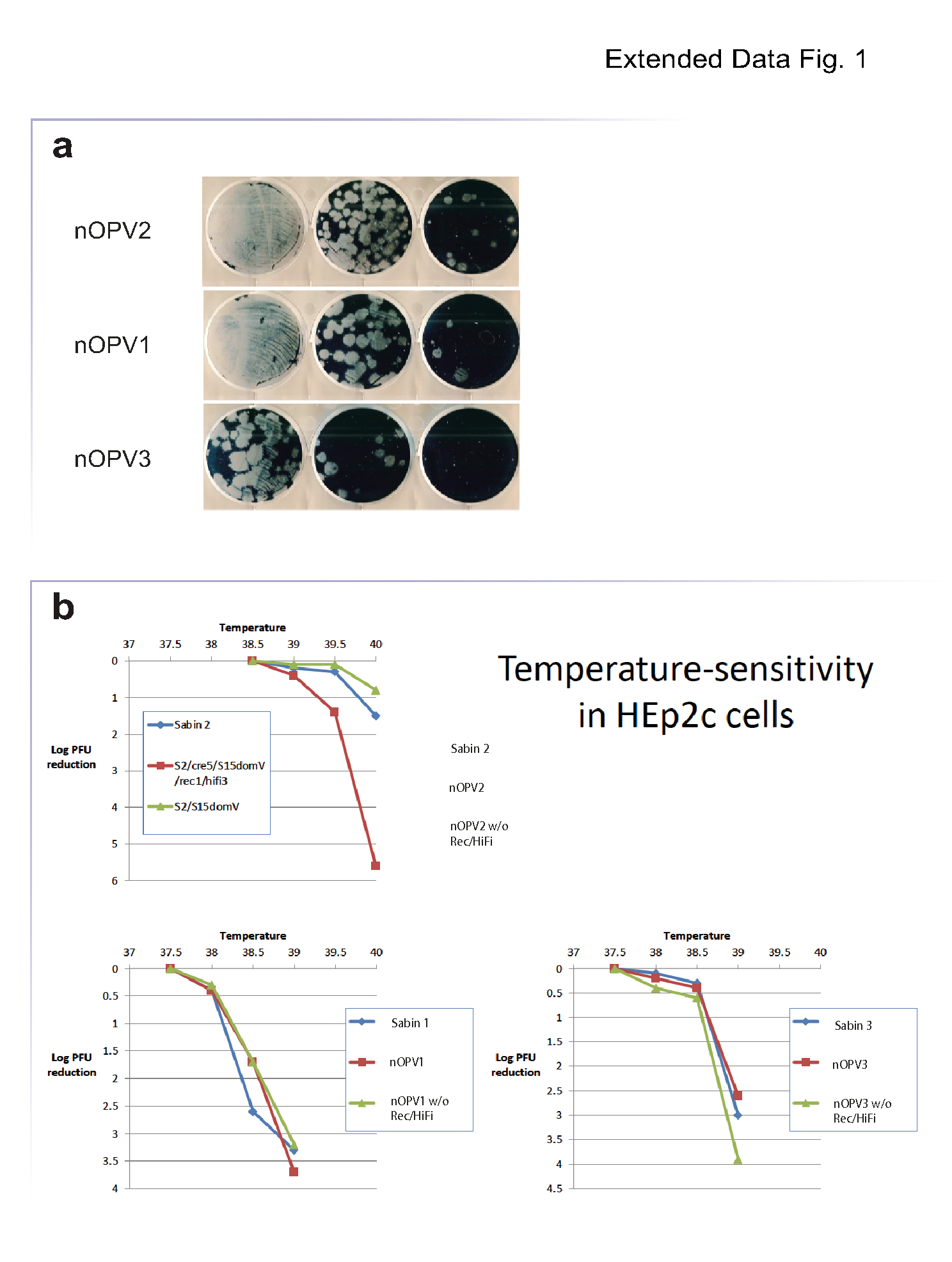
A critical aspect of any live-attenuated vaccine is the ability to grow to levels sufficient for production. Thus, we examined the fitness and replication properties of nOPV1 and nOPV3 carrying all the modifications described above (Fig. 1a) in Vero cells used for vaccine production35. The plaque size phenotypes of nOPV1 and nOPV3 were not significantly different than those of nOPV2 (Extended Data Fig. 1a). In addition, replication fitness was not reduced as shown by one-step growth analysis (Fig. 1b). In Vero cells at 33ºC, nOPV1 and nOPV3 replicated to higher titers than Sabin 1 and Sabin 3 with similar growth kinetics, but they maintained their temperature sensitivity at 37º and 39.5ºC as did the Sabin strains (Fig. 1c).
Importantly, because each modification in nOPV1 and nOPV3 (cre5, S15 domV, HiFi-D53N and Rec1-K38R) contributes independently to attenuation 22, their combination in the same virus vaccine strains creates a multilayer safety-net that reduces the likelihood of the new Sabin strains regaining fitness and neurovirulence. Accordingly, reversion to a pathogenic virus from the vaccine will require multiple independent mutation events, including several low-probability AU-to-GC double mutations within domV, a double recombination event in the 5′-UTR to replace S15 domV and preserve the cre5, and a reversion of the two polymerase modifications that do not reduce replication fitness. A critical feature of this virus vaccine design is that nOPV1 and nOPV3 maintain their replicative capacity in tissues without causing disease, which is essential to elicit a robust life-long immune response.
To examine the effects of these substitutions, we initially used a cell-culture assay that estimated temperature sensitivity, which is a characteristic correlating with attenuation4,21 . The temperature sensitivity assay indicated that replacing domV with S15 had little or no effect on Sabin 2 sensitivity at the tested temperatures (Extended Data Fig. 1a), whereas introduction of 3D polymerase modifications and the cre5 relocation produced viruses that were more temperature-sensitive than Sabin2 (Extended Data Fig. 1a). In contrast, nOPV1 and nOPV3 chimeric viruses were as thermosensitive as Sabin 1 and Sabin 3 with or without the cre5 relocation and 3D polymerase modifications (Extended Data Fig. 1b and 1c).
We determined the median paralysis dose (PD50) for each of these mutant viruses as described in the Methods section (Fig. 1d). To do this, we used a WHO-recognized mouse neurovirulence test (https://www.who.int/biologicals/vaccines/) that is used to test new lots of Sabin vaccines. The PD50 (log TCID50) was determined as 3.1, 5.9, and 3.9 for the Sabin 1, 2, and 3, respectively. All three new vaccine strains, nOPV1, nOPV2 and nOPV3 with PD50s of greater than 8, were more highly attenuated than Sabin 1, 2 and 3 (Fig. 1e).
To further examine neuroinvasion and neurovirulence, we inoculated susceptible mice intraperitoneally with various dosages of nOPV1-c1, nOPV3-c1, Sabin 1 and Sabin 3. After inoculation with the higher doses, no mice inoculated with Sabin 1 survived, and 60% of the animals inoculated with Sabin 3 also showed symptoms of severe paralysis (Extended Data Fig. 2b and d). In contrast, most mice survived inoculation with the highest doses of nOPV1 and nOPV3 (Extended Data Fig. 2c and e). Thus, in this mouse model as well, nOPV1 and nOPV3 are more attenuated than the original Sabin 1 and Sabin 3.
Genetic stability of nOPV1 and nOPV3
Next, we determined whether nOPV1 and nOPV3 are indeed more genetically stable than the current Sabin 1 and 3. Initially, we used a simple cell-culture evolution paradigm (Fig. 2a). The thermosensitive phenotypes of Sabin 1 and Sabin 3 are more pronounced in certain cell lines. For example, the serial passage of Sabin viruses at 37°C in Vero cells, but not in Hep2C, promotes the rapid incorporation of mutations that increase both replicative fitness and thermotolerance 36. These thermoresistant variants also display a significant increase in virulence4,21
We thus carried out serial passage experiments using large viral population sizes and low multiplicities of infection (m.o.i.) (106 particle forming units (PFUs) and m.o.i.= 0.1) that further ensured rapid evolution37. After 10 passages, we sequenced the virus population of both nOPV1 and nOPV3 and parental Sabin 1 and 3 using next-generation sequencing to determine variant frequency. While the control Sabin vaccine strains acquired the neurovirulence determinants within domV, nucleotides 480 and 525 (5 and 63% for Sabin 1, Fig. 2b and Extended Data Fig. 3a) and 472 (~95% for Sabin 3, Fig. 2c and Extended Data Fig. 3b), no mutations with frequency of greater than 1% were identified within domV nor for the other four modifications introduced originally into nOPV2 and hence present in nOPV1 and nOPV3 (Fig. 3). However, we observed accumulation of mutations, mostly in the 2A region, at frequencies of 4–75% in both Sabin strains and nOPV1 and nOPV3 populations (Fig. 2b and c; and Extended Data Fig. 3a and b). These 2A mutations are associated with cell-culture adaptation and have no effect on monkey neurovirulence38. We also observed mutations in positions leading to substitutions in VP1 and VP3, as well as silent mutations in both Sabin and nOPV strains (Fig. 2a and b).
We next examined virulence of nOPV1 and 3 by direct intraspinal inoculation of transgenic mice expressing the human poliovirus receptor. The PD50s for Sabin 1 and Sabin 3 in this model were determined as 3.1 ad 3.9 log10 CCID50 (50% cell-culture infective dose), respectively (Fig. 2d, P0). Strikingly, nOPV1 and nOPV3 induced no paralysis in any mouse even at the highest available dose up to ~8.8 log10 CCID50, indicating significant lower neurovirulence than the Sabin strains. We also examined neurovirulence of populations of Sabin and nOPV vaccine candidates at high-temperature growth selection in Vero cells (Fig. 2d, P10). Consistent with the observation that Sabin 1 and Sabin 3 accumulate mutations within domV, the high-temperature-adapted Sabin 1 and Sabin 3 viruses exhibited reduced PD50 of 2.2 and 1.7 log10 CCID50, respectively. On the other hand, nOPV strains remained attenuated after 10 passages with no measurable neurovirulence at the highest tested dose (Fig. 2d). Thus, both deep sequencing and mouse neurovirulence testing confirmed that the designed vaccine candidate nOPV1 and nOPV3 are more resistant to reversion to neurovirulence than the Sabin strains.
Characterizing the effect on neurovirulence of mutations identified in clinical trials
In the context of two similar but non-contemporaneous clinical trials, 18–22-week-old infants and 1–5-year-old children were immunized with nOPV2 or Sabin 2. Virus shed from vaccinees was isolated and nucleotide polymorphisms examined by next generation sequencing23,27,30,31,39,40. There is also considerable sequence data from Sabin 2 isolates shed by vaccinees and contacts. Given that nOPV1 and nOPV3 are derived from nOPV2, we used these data to examine the possibility that nOPV1 and nOPV3 may acquire such mutations to increase fitness and even neurovirulence.
Substitutions at several positions within the 5′UTR were present at significant frequencies in viruses excreted by several subjects after about 2 weeks of replication, including mutations 215C, 322A, 459C and 498C in domains II, III and IV within the internal ribosome entry site. Mutations 123C or 181G at the base of the 5′cre stem were also observed. These mutations increase the stability of the 5′cre by converting a U-G base pair near the foot of the structure into a C-G or creating a new C-G base-pair at the foot of the stem. Mutation 459C (nucleotide 398 in Sabin2) was identified as a secondary adaptive mutation in cVDPV2 isolates and vaccinee isolates4. Mutations in capsid protein VP1 which increase neurovirulence (VP1-I143T/V) also were observed. The frequency of these mutations varied between 7 to 70% from individual to individual. However, only a small proportion of shed viruses carried all these mutations. Viruses that had fixed all three modifications, 123C or 181G, 459C, and I143T/V, were only found in samples taken several weeks post-vaccination 23,39,41.
To determine if the mutations observed in the virus shed from vaccinees could increase their fitness and virulence, we engineered each of them, in different combinations, into the infectious cDNA clones of nOPV1 and nOPV3. We introduced single or multiple substitutions 123C and 181G, 215C, 322A, 459C, and 498C. We replaced the capsid coding region with that derived from wild-type Mahoney type 1 and Leon type 3. We also generated nOPV1 and nOPV3 carrying six mutations within the 5′UTR (5′rev) (Fig. 3). Furthermore, we combined some of the 5′UTR mutations with the wild-type capsid replacement. Although we did not attempt to analyze all possible combinations or evolutionary trajectories, this extensive analysis provided a rich set of information that can be useful to evaluate the probability of nOPV1 and nOPV3 to evolve into neurovirulent forms after circulation in humans (Fig. 3). For example, the reconstructed 5′rev/MahP1 and 5′rev/LeonP1, containing all 5′UTR mutations and replacement of P1 region represent the “worst-case” scenario in which many of the mutations that were selected in shed viruses after many weeks of replication, had accumulated to 100% in a single genome, imparting the maximum increase of fitness23,39,41. As comparisons, we used Sabin1/480A and Sabin3/472C, which are the de facto cases when Sabin 1 and Sabin 3 are employed (i.e., a revertant virus excreted by most vaccinees within 1 or 2 weeks of vaccination). When neurovirulence was evaluated by intra-spinal inoculation of susceptible mice, 480A in Sabin 1 or 472C in Sabin 3 produced neurovirulent viruses with a PD50s of 1.6 and 1.3 (log10 TCID50), respectively. The mutant 5′rev/MahP1 and 5′rev/LeonP1 had PD50s of 1.5 and 3.9. Thus, even in the unlikely event that nOPV3 accumulated all changes in one genome, the new strain would be at around 400 times more attenuated than the single-mutation Sabin 3 revertant (Sabin3/472C), which is produced almost universally days after vaccination. For Sabin 1, we find that the acquisition of all the observed mutations will render a virus that is similar to reverted Sabin1/480A, but to acquire that level of neurovirulence, the virus would need to acquire six independent 5′UTR mutations and multiple mutations in the P1 region. Importantly, all the intermediate viruses with fewer mutations are much less virulent than Sabin strains with a single mutation within domV (Fig. 3).
Antigenic and immunogenic characteristics of nOPV1 and 3
Because the nOPV1 and nOPV3 strains are intended to elicit immune protection against virulent poliovirus strains, we analyzed the antigenic structure of the vaccine candidate in an ELISA assay similar to that used to measure the D-antigen content of poliovirus vaccines. A panel of monoclonal antibodies (MAbs), specific for Sabin1 native conformations of antigenic sites 1 (MAb 955), 2 (MAbs 237), 3 (MAb 424), and 4 (MAbs 234), were used as primary antibodies42. We also examined Sabin3 native conformations of antigenic sites, using monoclonal antibodies directed against site 1 (MAb 520), 2 (MAbs 877), 3 (MAb 883), and 4 (MAbs 1281) 42,43. The Sabin 1 and Sabin 3 strains, as well as IPV1 and IPV3, were used as a reference in each experiment. Differences in antigenicity from Sabin references were not noted at any of the four sites tested for type 1 and type 3 polioviruses (Extended Data Fig. 4a and b).
We then evaluated nOPV1 and nOPV3 immunogenicity in a model requiring efficient replication for induction of antibody responses. Susceptible transgenic mice were infected intraperitoneally with dilutions of Sabin 1, Sabin 3, nOPV1 or nOPV3, and serum samples were collected at 21 days post-inoculation and tested by neutralization (NT) assay to determine antibody titers (Fig. 4a). Inoculation with Sabin 1 tended to induce higher geometric mean titers of neutralizing antibodies, particularly at low doses (Fig. 4b). However, the difference between antibody titers induced by Sabin 1 and nOPV1 at the tested doses and at the tested sample size did not reach statistical significance, except for low doses (e.g., 104 TCID50) (Fig. 4b, two-tailed Mann-Whitney U test). Similarly, we observed no significant difference in antibody titer in animals inoculated with Sabin 3 and nOPV3 (Fig. 4c, two-tailed Mann-Whitney U test). Importantly, Sabin 1, Sabin 3, nOPV1 and nOPV3 had similar seroconversion rates, and most vaccinated mice generated neutralizing antibodies at the tested doses with a single immunization (Fig. 4b and c, numbers on the top of the graphic). Hence the immunogenicity of nOPV1 and nOPV3 were not significantly inferior to that of Sabin strains.
Finally, we compared the immunogenicity of the new nOPV vaccine candidates to Sabin strains in bivalent and trivalent formulations. Each Sabin strain (Sabin 1, Sabin 2 and Sabin 3) and nOPV1, nOPV2 and nOPV3 were mixed in a 1:1:1 proportion of each virus. Mice were inoculated intraperitoneally with 3x106 TCID50 of each virus of the bivalent and trivalent formulation. We observed no significant difference in antibody titer in animals inoculated with Sabin and nOPV bivalent or trivalent formulation (Fig. 5a and b, two-tailed Mann-Whitney U test). Furthermore, as with the monovalent formulation, we observed similar seroconversion rates for Sabin and nOPV bivalent and trivalent formulations (Fig. 5a and b, numbers on the top of the graphic).
We also assessed the correlation between seroconversion and protection in this animal model. Vaccinated mice were challenged with highly pathogenic polioviruses (Mahoney type 1, MEF type 2, and Leon type 3) 4 weeks after vaccination. We observed similar levels of protection for Sabin and nOPV vaccinated groups after a single vaccination dose (Fig. 5c and d). Interestingly, consistent with the lower PD50s and rates of reversion observed in Sabin strains, several mice immunized with Sabin strains became paralyzed or died (Fig 5a and b, asterisks), but none of the nOPV vaccinated animals presented any signal of disease.
{kind=link}
{kind=link}
{kind=link}
{kind=link}