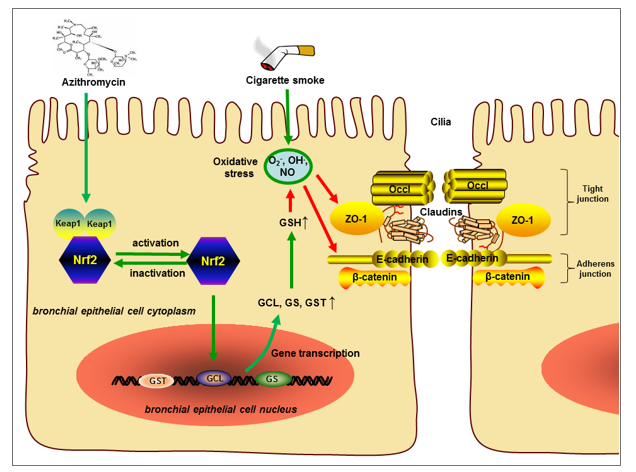
In our present study, we found that AZI not only possessed anti-inflammatory and anti-oxidative properties, but also ameliorated airway epithelial barrier dysfunction by improving TEER and apical junctional complexes. Our study also revealed that AZI significantly up-regulated the expression of Nrf2 to promote GSH metabolism, highlighting the novel role of Nrf2/GCL/GSH signaling pathway in maintaining airway epithelial barrier function. These results demonstrated that AZI prevented CS-induced airway epithelial barrier dysfunction through Nrf2/GCL/GSH signaling pathway.
The bronchial epithelium is responsible for maintaining the airway homeostasis of respiratory system. Destruction of airway barrier integrity exposes sub-epithelial layer to inhaled particles, triggering airway inflammation and immune responses, indicating that airway epithelial barrier dysfunction is closely related with respiratory diseases [5, 9]. It has been confirmed that smoking disrupted apical junctions of airway epithelium, and the reduction of apical junction genes has been observed in the lung tissues of COPD patients [31]. In addition, TJ proteins were also significantly suppressed in lung tissue of patients with end-stage COPD and in air-liquid interface differentiated epithelial cells from these patients [13]. Similarly, our experiments in vitro revealed that CSE exposure caused the degradations of TJ protein ZO-1 and AJ protein E-cadherin with subsequent TEER decline, which eventually leads to airway epithelial barrier dysfunction. It was well known that structural and subsequent functional destruction of epithelial barrier is a typical feature of chronic airway inflammation. Many innate and adaptive immune mediators that may be up-regulated after long-time cigarette smoking, including cytokines, chemokines and apoptosis factors, could regulate the airway epithelium barrier function [5]. As found in our study, CS-increased secretions of pro-inflammatory cytokines and airway epithelium cell apoptosis in vitro and in vivo, which was consistent with above reports.
AZI is commonly indicated for the treatment of respiratory bacterial infection, and exerts immunomodulatory activities in chronic inflammatory disorders, such as COPD [32–34]. Clinically, in patients with severe COPD, continuous therapy of AZI combined with nebulized colistin dramatically prevented the exacerbations of COPD [35]. Preventive administration of AZI reduced the frequency of acute exacerbation and improved the quality of life in COPD patients [20]. However, the underlying mechanism is not completely clear. Here, our study provided evidence that AZI treatment counteracted the CSE-induced TEER reduction and disruption of ZO-1 and E-cadherin, along with the inhibition of inflammatory response and apoptosis in vitro and in vivo. Consistently, a recent study revealed that AZI treatment substantially enhanced epidermal characteristics partially by up-regulation of tight junction proteins in bronchial epithelial cells [36]. In addition, pretreatment of AZI inhibited the secretions of IL-6 and IL-8 in CS-exposed bronchial epithelial cells, suggesting that AZI may be beneficial for smoking-induced airway epithelial barrier dysfunction [37].
Metabolomics is considered as a promising method to accurately determine all low molecular weight metabolites of an organism and reveal its biology and response to pathophysiological stimuli [38]. The results of metabolomics usually contain a continuous stream of high-content information, which will essentially help to understand the differentiating metabolite profiles from a global perspective [39]. Therefore, metabolomics has been widely applied in various fields, such as drug toxicity, disease diagnosis, and pharmacodynamic study [40–42]. To elucidate the mechanisms though which AZI ameliorated CSE-induced airway epithelial barrier dysfunction, we performed metabolomics profiling. Interestingly, our experimental results showed that metabolite set enrichment analysis revealed pathways upregulated by AZI treatment, including GSH metabolism. GSH, the most abundant non-protein thiol compounds in mammalian tissues and cells, is known as the most important endogenous molecule to resist oxidative stress, detoxify xenobiotics and regulate cell proliferation, apoptosis, immune function, and fibrogenesis [43]. Due to the central role of GSH in maintaining cellular redox homeostasis, it is absolutely necessary for a series of biochemical reactions to protect airway epithelial cells from CS-induced oxidative stress [44]. It has been known that CS-induced oxidative stress weakened GSH levels in airway epithelium and disrupted tight junctions, epithelial barrier integrity, finally leading to the impairment of epithelial barrier function [45, 46]. Herein, our study showed that the major molecules or enzymes involved in GSH synthesis and metabolism pathway, including GSH, GCL, GS, GST, L-Glutamic acid and pyroglutamic acid, were all significantly decreased by CSE exposure, while these molecules or enzyme activity were prominently restored by AZI treatment. In fact, AZI has already been reported to suppress ROS release in epithelial cells and prevent oxidative damage in macrophages harvested from 8 transplant recipients [47], indicating the antioxidant potentials of AZI. For the first time to our knowledge, our experimental results highlighted a role for AZI promoting glutathione metabolism in the lungs of CS exposure.
Oxidative stress is highly correlated with the impairment of glutathione metabolism in COPD pathogenesis [48]. GSH synthesis and metabolism are mainly determined by GCL, GS, GST, which are directly regulated by Nrf2 transactivation [49, 50]. Nrf2, a transcription factor significantly expressed in airway epithelial cells, is known to regulate antioxidant and cytoprotective genes through activating antioxidant response elements, showing protective effects on airway epithelium [51]. It has been confirmed that Nrf2 and some of its target genes constituted a protective signaling pathway against oxidative stress and inflammation in COPD development [52, 53]. And the deficiency of Nrf2 contributes to the pathogenesis of COPD, accompanied with dysregulation of GSH metabolism [49, 51, 54, 55]. A recent observational longitudinal study revealed that GSH was significantly reduced in the blood samples of COPD patients, moreover the expression of Nrf2 in PBMCs were significantly down-regulated in COPD patients at follow-up compared with non-COPD patients [56]. Also, it was reported that activating Nrf2/GCL/GSH antioxidant signaling pathway by quercetin could attenuate toosendanin-induced oxidative stress to prevent hepatotoxicity [57]. As expected, our results demonstrated that AZI treatment significantly prevented Nrf2 suppression which was induced by CS exposure in vitro and in vivo. Moreover, our experiments further verified the role of Nrf2 in AZI treatment by using Nrf2-shRNA, Nrf2 agonists, antioxidant and Nrf2 knockout mice, confirming that the protective effects of AZI on CS-induced airway epithelial barrier dysfunction primarily depends on the activation of Nrf2/GCL/GSH signaling pathway.
It is also noteworthy that not only AZI, but some other macrolides (e.g. erythromycin, clarithromycin) have also been reported to prevent COPD exacerbations and improve patient quality of life and symptoms [58]. Thus, to further explore whether the protection of airway epithelial barrier is the common pharmacological activity of macrolides, we have repeated the in vitro experiments using erythromycin (EI, 14-membered ring macrolide) and spiramycin (SPI, 16-membered ring macrolide). Data in Fig. 8 revealed that EI could also protect the airway epithelial barrier against CSE and its molecular mechanism was similar to that of AZI, while SPI had no similar pharmacological activity and regulatory effect. Nevertheless, there are some limitations in this study: 1) We only investigated the preventive effect of AZI, but its clinical therapeutic effect on protecting airway epithelial barrier is still worthy of further verification; 2) We have already found that Nrf2 is the key mediator in the protective effect of AZI on airway epithelial barrier, but how AZI activates Nrf2 is still unknown, which is our next research objective; 3) Different pharmacological effects of macrolides on airway epithelial barrier may be related to the difference of structural characteristics, which requires molecular docking and structure modeling methods to further clarify the underlying mechanisms.
{kind=link}