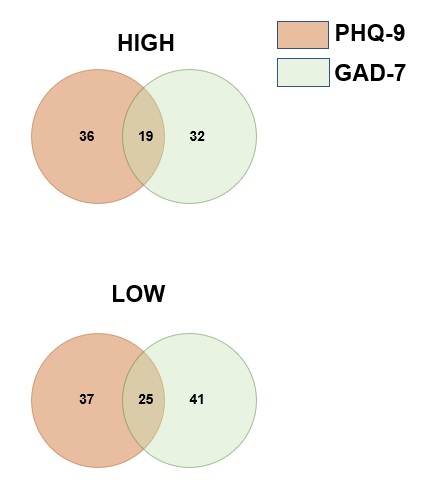
Being diagnosed with a tumor and underwent surgery, especially brain surgery, may bring major negative impact on one’s life. Some of the patients might just meet transient or mild negative mood about the diagnosis or treatment, but some may suffer persistent negative mood in a moderate or severe level, could then develop into a multifaceted anhedonic state, and impaired cognitive function, which could be defined as depressive/anxiety disorders [24]. It had been showed by many researches the certain correlation of tumor with depressive/anxiety disorders, and some of these researches are focusing on the influence of depressive/anxiety on the outcome of tumor patients [31]. But none of these researches provided any information about the potential mechanism of depression/anxiety disorder on the prognosis of tumor patients. Measured with the widely used scale PHQ-9 and GAD-7 questionnaires, we firstly screened out the psychiatric condition of 73 GBM patients in our department. We found that those GBM patients with higher PHQ-9 and GAD-7 score have significantly worse prognosis than their lower scored counterparts. It seems the case that glioblastoma and depressive/anxiety disorder share some common etiological factors, such as inflammation [32, 33], neurotransmitter metabolism [34], glutamine [35, 36], neural plasticity [37, 38], etc. Nevertheless, none of these factors were confirmed by further researches. With the rapid development of human genome project and the cancer genome atlas system, it is now possible for us to start a genetic-level analysis to evaluate genetic alternations across diseases. As diseases with multiple genetic abnormalities, there should be a possibility that there are some genetic alternations overlapped in the brain which may not only result in susceptibility to depression and anxiety, but also in the malignant progression of GBM which result in worse prognosis. As a result, the bioinformatic mining were conducted.
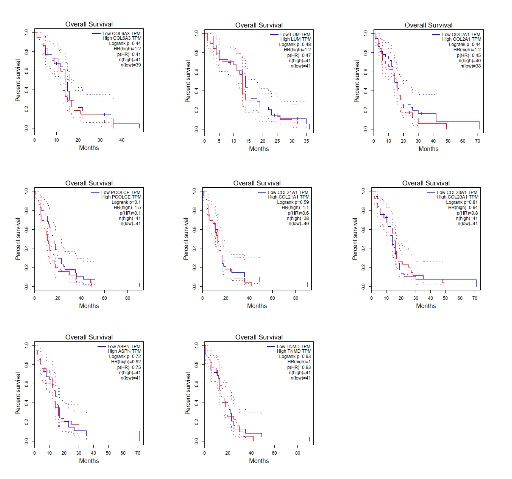
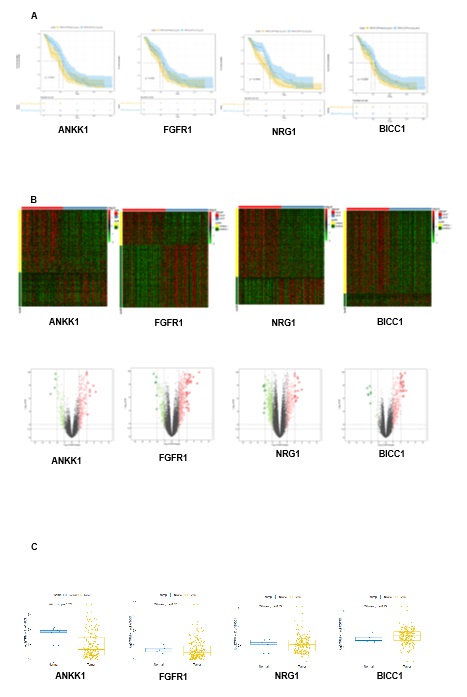
Using a series of bioinformatic methods, six genes related to depressive/anxiety disorders which have negative impact on the prognosis of GBM were screened out. And among these genes, LTBP1 was differentially expressed between tumor and normal tissue. LTBP1, as an important member of the latent TGF-β1-binding protein family, has been shown to bind to the latent form of TGF-β and escort it during secretion, thus enhancing TGF-β bioavailability [25]. Researchers have found that LTBP1 could play a pivotal role during the processes of psychomotor retardance and disorders [39]. And LTBP1 and its target TGF-β1 were also involved in Alzheimer disease and depression [28]. On the other side, it was also reported that LTBP1 was closely involved in the processes of glioma [29, 40]. However, there is neither evidence that can provide the exact mechanism of LTBP1 on depressive/anxiety disorder as well as GBM, nor any researches focusing on the connection linking depressive/anxiety disorders and the prognosis of GBM. We discovered that the expression of LTBP1 was positively correlated with the PHQ-9 and GAD-7 score among these GBM patients. This encouraged us to further evaluate the function of LTBP1 and possibly find a potential bridge built by LTBP1 that could link depressive/anxiety and GBM.
We firstly confirmed the LTBP1 negative influence of LTBP1 on the outcome of gliomas with the data from Chinese Glioma Genome Atlas. We then found that the primary GBM cell with higher expression of LTBP1 have stronger proliferation and migration capacity than those with lower expression of LTBP1. These results indicated the direct function of LTBP1 on GBM. We further conducted the functional enrichment analysis and the protein-protein interaction of the differentially expressed genes regulated by LTBP1. To our astonishment, the findings revealed that the enriched functions and pathways were mostly related to extracellular matrix (ECM). And hub genes closely related to collagen formation and ECM, such as COL1A2, COL10A, etc. which were also key indicators on GBM outcome, were also differentially expressed between high and low PHQ-9 and GAD-7 groups in our research.
Extracellular matrix seems to be a probable mechanism by which we could link the depressive/anxiety disorder and worse outcome of GBM. In terms of tumor, including GBM, the ECM regulates tissue development and homeostasis, and its dysregulation contributes to neoplastic progression [41]. The cancer-associated ECM is not only an integral feature of a tumor but also actively contributes to its histopathology and behavior [42]. Researches had discovered that ECM-associated alternations could overwhelmingly influence the capabilities of human tumors including sustained proliferation [43], evasion of growth suppression [30], death resistance [44], replicative immortality [45], induced angiogenesis [46], initiation of invasion [47], dysregulation of cellular energetics [48], avoidance of immune destruction [49] and chronic inflammation [50]. During the tumorigenesis and development gliomas, significant alterations of the ECM in brain tissues could be triggered by glioma tissue. Such changes include altered synthesis of the components by the tumor cells, extensive degradation of the ECM at the invasive front of the tumor, as well as an elevated level of synthesis of ECM components by normal tissues in the vicinity of the invading tumor. Moreover, it was revealed that glioma cells had the ability to actively migrate using blood vessels or axons as guide paths due to interaction with the ECM. Additionally, glioma cells can secrete their own ECM components, including HA, brevikan, tenascin C and thrombospondin, as well as fibronectin, which are actively expressed in the ECM of the developing nervous system along cell migration paths. [51, 52]. This could partly explain that the in vitro GBM cell could also proliferate significantly different in our experiment. On the other hand, the function of ECM was also significant factor on the process of depressive/anxiety disorders. It is discovered that ECM alterations in the cognitive component was associated with depressive-like behavior [53, 54]. And ECM markers such as MMP9 and sICAM1 were also reported to deficit cognitive function in human brain which may result in serious psychiatric disorders such as depressive disorders and bipolar disorders [55].
To our knowledge, there are many possibilities that the alternation of ECM by differentially expression of LTBP1 in brain may link depressive/anxiety disorders and glioblastoma (Fig. 10). Firstly, Brain neural tissue functions as a dynamic network—beneficial synaptic connections need to be maintained, and other reconstructed to match changing input stimuli. Cell–cell interactions in the brain, similarly to other tissues, are based on direct contacts via cadherins and signaling receptors, as well as cell–matrix interactions with the ECM [56]. Differentially expressed LTBP1 could change the components of ECM and stabilization of network, alter the neural plasticity and change the adhesion and interaction among of brain, which may result in susceptibility of neurological and psychological disorders, including depressive/anxiety disorders. Similarly, such alternation could also trigger the aggressiveness and migration of GBM cells and result in worse prognosis [57]. Secondly, LTBP-1 targets latent TGF-β1 to the ECM by interacting with different proteins including fibronectin and fibrillin, generating deposits of latent TGF-β1 accessible for cell-mediated activation and regulating cancer cell proliferation and immunity [58]. Immunology homeostasis instabilities may cause deficits of cognitive and memory in human brain, which could be a potential cause of depressive and anxiety disorders[59]. On the other side, as important component of neural microenvironment, ECM could regulate the neural inflammation in central nervous system[60]. Inflammation is believed to be a typical pathology changes in depressive and anxiety disorder [61], and chronic inflammation may also result in the epithelial mesenchymal transformation (EMT) of glioblastoma, and in turn cause worse outcome in GBM patients [62]. The changing of ECM by LTBP1 could function as an indicator of depressive/anxiety disorder and GBM by regulating inflammatory responses. And in order to confirm the case, we evaluated the expression of epithelial and mesenchymal markers in high and low LTBP1 group and found a higher expression of mesenchymal markers such as vimentin and CD44, as well as lower expression of typical epithelial markers such as E-cadherin, in high LTBP1 group. As for the reason why no expression differences of SOX1 was observed between the two group, we suppose that SOX1 is a transcription factor that expressed almost in all tissue and cells with high proliferative rate and stemness. All subtype of GBM have a relatively high expression of SOX1, so no significantly despite slightly difference could be observed between high and low LTBP1 groups. But these findings are enough to prove the evidences that there were more cells classified as mesenchymal subgroup between primary GBM cells from high and low LTBP1 groups, namely those with high and low PHQ-9 and GAD-7 scores, which may result in the significant difference of proliferation and migration rate. However, due to the lack of animal model that could mimic both depressive/anxiety as well as GBM, we could not provide any in vivo experiments that directly prove the evidence of the interaction between depressive/anxiety disorders and GBM. Our speculation was based on only bioinformatic and cellular experimental results, and further researches were still needed to elucidate the exact mechanism of this phenomenon.
{kind=link}
{kind=link}
{kind=link}