3.1 Gene expression analysis of ZMYND11
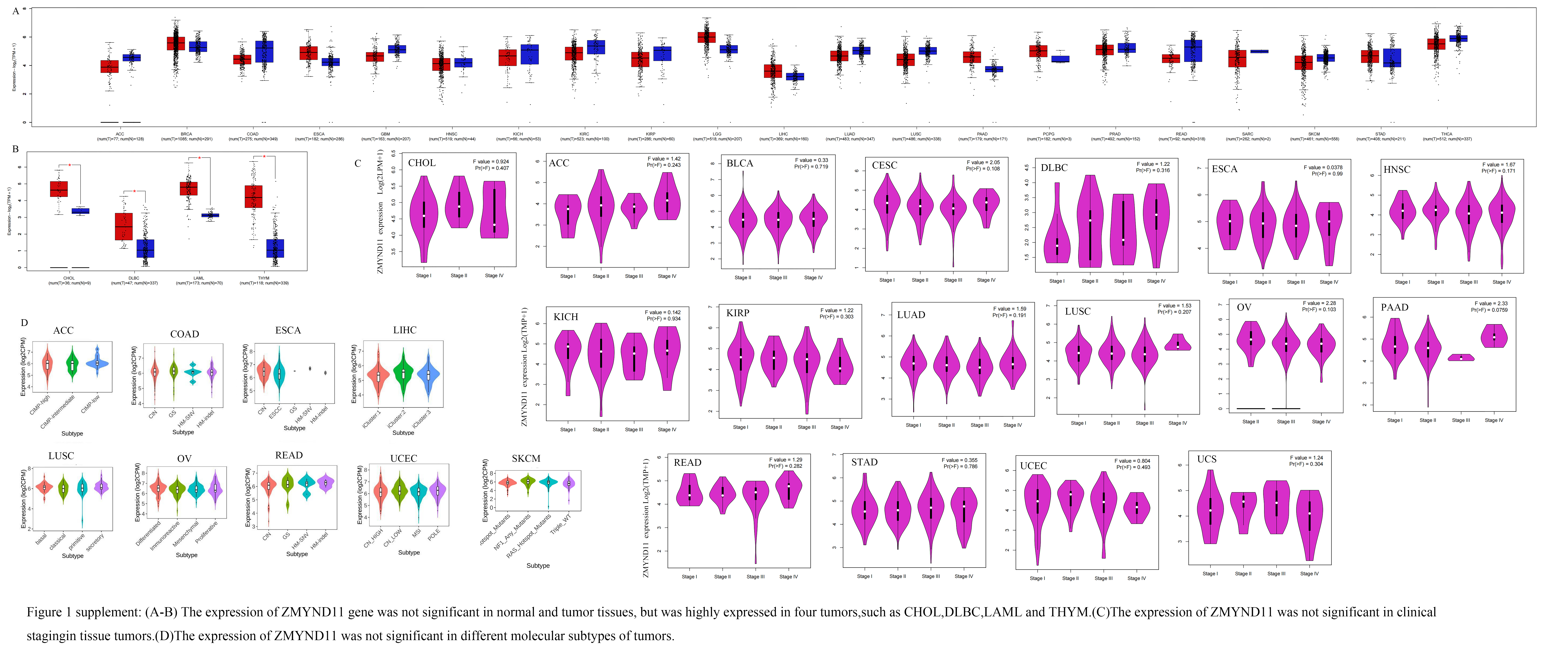
First, we used the TIMER2 database to identify the expression states of the ZMYND11 in various carcinoma types from TCGA. Figure 1A dispiays that in control tissues, the expression of ZMYND11 was reduced in BLCA, BRCA, CESC, GBM, KICH, KIRC, KIRP, LUAD, LUSC, PRAD, READ, THCA, and UCEC(P < 0.05). And later in the common organization from the GTEx dataset as a comparison, the expression ZMYND11 was reduced only in BLCA, CESC, OV, TGCT, UCEC ,UCS (Fig. 1B, P < 0.01), and in CHOL, DLBC, LAML, THYM was highly expressed (shown in Supplementary Fig. 1B, P < 0.01). We did not observe remarkable distinctions for the other neoplasms, such as ACC, LUAD, PAAD, and others (Supplementary Fig. 1A).
In the CPTAC database, we found that the aggregate protein of ZMYND11 was less expressed in original oncologic tissues of COAD, KIRC, UCEC, LUAD, PAAD, and HNSC than in natural organizations. However, it was moderately expressed in BRCA and GBM (Fig. 1C, p < 0.05). We then analyzed the ZMYND11 mRNA expression level at various clinical stages and subtypes. The association between ZMYND11 expression and stage of disease was observed by the module of HEPIA2 "pathological staging map," and significant differences were found between the various phases of the BRCA, COAD, KIRC, SKCM, THCA, and LIHC (Fig. 1D, p < 0.05). In the expression of ZMYND11 at the various molecular subtypes of a STAD, HNSC, KIRP, BRCA, GBM, LGG, PCPG and PRAD has a remarkable significant differences. (Fig. 1E, p < 0.05). There was no connection between ZNYND11 expression and carcinoma grade or subtypes of molecules in other cancers (Supplementary Figs. 1D-E).
Finally, we concluded that ZMYND11 is lowly expressed in BLCA, BRCA, CESC, GBM, KICH, KIRC, KIRP,LUAD, LUSC, PRAD, READ, THCA and UCEC in TCGA. Furthermore ,the expression of ZMYND11 gene was reduced in BLCA, CESC, OV, TGCT, UCEC and UCS in TCGA and GTEx data .
3.2 Prognostic analysis of survival
Based on the use of the website ( https://www.aclbi.com/static/index.html#/), We first analysed the differences in expression and prognosis between the 33 types of tumors in TCGA using gene expression data. Using one-way survival analysis, we investigated the forest plot of the ZMYND11 expresssion in 33 tumors Overall survival (Fig. 2A and Supplementary table 1). Furtherly we analyzed the differences between the gene values that were associated with the 33 types of TCGA cancer and their Disease Free Survival (Fig. 2B and Supplementary table 1).And this results indicated that ZMYND11 is a good prognostic factor in ACC,KIRC,LGG,LUAD,MESO,PRAD,READ and THCA. For the 33 tumors in TCGA, the differences in gene expression values and the prognostic intervals for Disease-free survival were considered; this result is displayed in Fig. 2C and Supplementary table 1. We also performed an additional analysis of the differences among the gene expression values from the 33 types of tumors in TCGA and their Disease specific survival. Considering possible non-tumor mortality factors during patient follow-up. The results are presented in Fig. 2D and Supplementary table 1.
We distinguished between tumor types with the high expressing and the low expressing of ZMYND11 according to their expression levels, and the TCGA dataset was used to identify the relationship between ZMYND11 expression in various tumor patients and their prognosis. Upon Kaplan-Meier survival curve analysis, the overexpression of ZMYND11 was prognostically more relevant to OS and RFS in patients with low expression of ZMYND11 KIRC,LGG,LUAD, MESO,PRAD,READ and THCA( Fig. 3A-B).
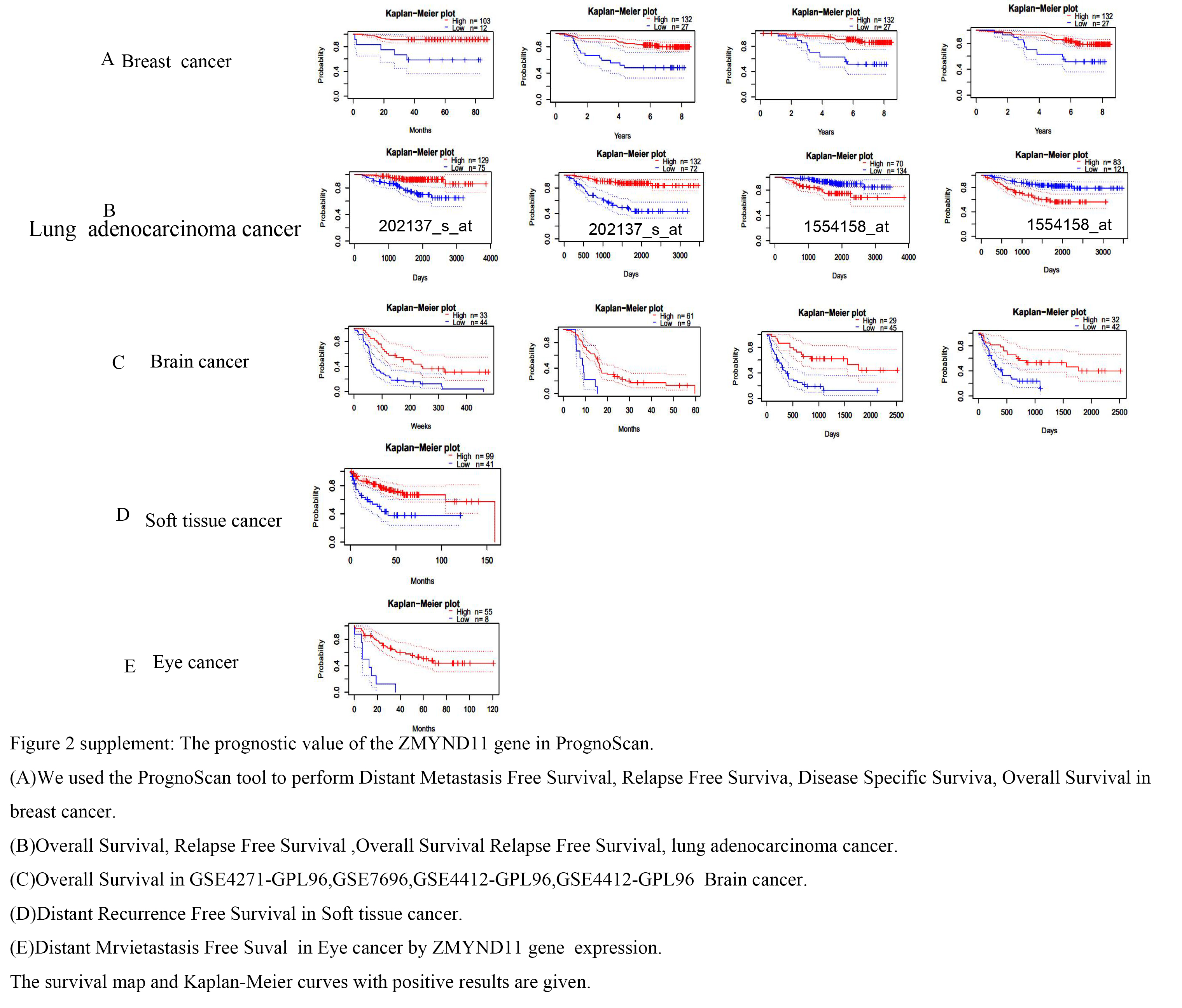
Furthermore, the PrognoScan database in the website (http://dna00.bio.kyutech.ac.jp/PrognoScan/index.html), which is based on the GEO database, was subjected to survival prognosis analysis for different tumors. We revealed that the high expression of ZMYND11 in the BRCA datasets GSE19615 (DMFS), GSE1456-GPL96 (RFS), GSE1456-GPL96 (DSS), and GSE1456-GPL96 (OS) had a comparatively favorable prognosis (Supplementary Fig. 2A). Probe ID 202137_s_ at GSE31210 (OS) and GSE31210 (RFS) had a high prognosis compared to lung cancer; however, in the lung cancer probe Probe ID 1554158_at GSE31210 (OS), GSE31210 (RFS) with low expression was relatively good (Supplementary Fig. 2B). It was also subsequently observed that the high expression of ZMYND11 in brain cancer GSE4271-GPL96 (OS), GSE7696 (OS), and GSE4412-GPL96 (OS) was associated with a better prognosis (Supplementary Fig. 2C). High expression in the soft tissue cancer dataset GSE30929 (DRFS) and eye cancer dataset GSE22138 (DMFS) had a superior prognosis (Supplementary Fig. 2D-E).
Overall, Based on the one-way survival analysis, Our results demonstrated that ZMYND11 could function and have a favorable overall survival prognosis in KIRC,LGG,MESO,LUAD,PRAD,READ and THCA. And With PrognoScan Kaplan-Meier survival curve analysis high expression of ZMYND11 offers a more favourable prognosis in Breast cancer ,Lung adencarcinoma cancer,Brain cancer ,Soft tissue cancer ,Eye cancer. In the end, Kaplan-Meier survival curve analysis depicted that high expression of ZMYND11 offers a more favorable prognosis in KIRC, LGG, MESO,, LUAD, PRAD,READ ,THCA,BRCA, SARC and UVM.
3.3 Analysis of mutations the ZMYND11 gene
Genetic and circumstantial elements are responsible for the development of genomic damage. Cancer is a complicated genetic disease linked with genomic instability,and genetic, and environmental components are important factors in regulating cancer development [19]. Genetic alterations in ZMYND11 were further explored using the cBioPortal (Fig. 4A).We found significantly higher mutation frequencies in BLCA, UCEC and OV tumors with low ZMYND11 expression, whereas mutation frequencies were very high in SARC and STAD. As you can see, among the mutations in the cBioportal database, we discovered that the missense mutations accounted for the primary types of ZMYND11 protein. Furthermore, we analyzed the COSMIC database(https://cancer.sanger.ac.uk/cosmic), a database of cancer somatic mutations covering various somatic mutation mechanisms that cause human cancers (1.40%), missense substitutions (23.31%), synonymous substitutions (6.62%),frameshift insertions (0.13%), frameshift deletions (0.38%), and others (6.11%). In total, 785 samples were identified; as in the cBioPortal database, missense mutations accounted for the significant mutations(Fig. 4B).Somatic mutations disrupt normal surveillance mechanisms and the emergence of other properties that favor unlimited cell proliferation and free invasion, thus transforming the cells into malignant. Mutations in ZMYND11 somatic cells might be involved in the tumorigenic process. In UCEC, three K441N/T mutations were found. In the PWWP structure of ZMYND11, alterations at the N304Kfs*33 and K345Rfs*43 sites were primarily found in one prostate cancer and one colorectal cancer. The N271Tfs*18 site was found in the importance of the Bromo and PWWP structural domains in invasive breast cancer. This may induce a gene mobility mutation, where asparagine translates to lysine at site 304 of the ZMYND11 protein, lysine translates to arginine at site 345, and asparagine translates to threonine at site 271, with a change in protein site translation that subsequently triggers truncation of the ZMYND11 protein. Gene fusions also play a paramount role in tumors. The current research indicated that a chromosomal translocation ZMYND11-MBTD1 gene fusion was diagnosed in human AML patients with acute myeloid leukaemia [8]. Moreover, we identified seven genes fused to ZMYND11-SORBS2,FAM208B-ZMYND11 in glioma and astrocytic glioma; ZMYND11-CDH18 gene fusion in bladder urothelial carcinoma; ZMYND11-LYZL1, RSU1-ZMYND11gene fusion in prostate adenocarcinoma; ZMYND11-PCDH15 gene fusion in stomach adenocarcinoma; CELF2-ZMYND11 gene fusion in Colorectal Adenocarcinomagene fusion(Fig. 4C). The fusion of genes may promote the high expression of some oncogenes. In human AML, the ZMYND11-MBT4 gene fusion preserves hyper-expression of leukemia-promoting genes involving Hoxa, Meis1, Myb, Myc, and Sox4 [8]. In the analysis of mutations in the ZMYND11 gene, it was found that mutations in the ZMYND11 gene also affected altered mutation frequencies in other genes with DIP2C, TUBB8, LARP4B, TP53, ADARB2, DIP2C-AS1, GTPBP4, WDR37, IDI1 and IDI2 genes having high mutation rate compared to ZMYND11 without mutations. the mutation rate was increased (Fig. 4E). Subsequently, we addressed the potential relevance of ZMYND11 gene variants to the clinical survivor prognosis of distinct categories of these cases. As illustrated in (Fig. 4F), Brain Lower Grade Glioma, brain lower-grade glioma cases without alterations had a better prognosis for overall survival (P = 1.7 e-3), disease-specific survival (P = 0.03) and progression-free survival (P = 2.9 e-3) compared to cases without ZMYND11 gene alterations, but not for disease (P = 0.846). This is consistent with our previous analysis of good survival of high-expression ZMYND11 in LGG.
In addition, based on the the website ( https://www.aclbi.com/static/index.html#/). We are currrently analyzing the link between ZMYND11 expression on the one hand and tumor mutation burden/microsatellite instability (TMB/MSI). (Fig. 5A and Supplementary table 1)There was a unique positive correlation between the expression of ZMYND11 and TMB in BRCA (P = 0.00047), DLBC (P = 0.042), KIRC (P = 0.0035), LGG (P = 4.63E-09), LIHC (P = 0.03), PAAD (P = 0.03), PRAD (P = 0.002), SRAC (P = 0.003), THCA (P = 8.06E-07); however, ZMYND11 gene expression demonstrated a negative correlation with TMB in THYM (P = 2.13E-07) and SKCM (P = 0.004)tumors( Fig. 5B and Supplementary table 1). It depicts that the expression of ZMYND11 gene was remarkably more positively related to the MSI in COAD(P = 0.008), LUSC(P = 0.03), UCEC(P = 0.02), and READ (P = 0.0003), however, there was a significant negative correlation with MSI in DLBC (P = 1.06E-05) and PRAD (P = 0.0005).
3.4 DNA methylation analysis data
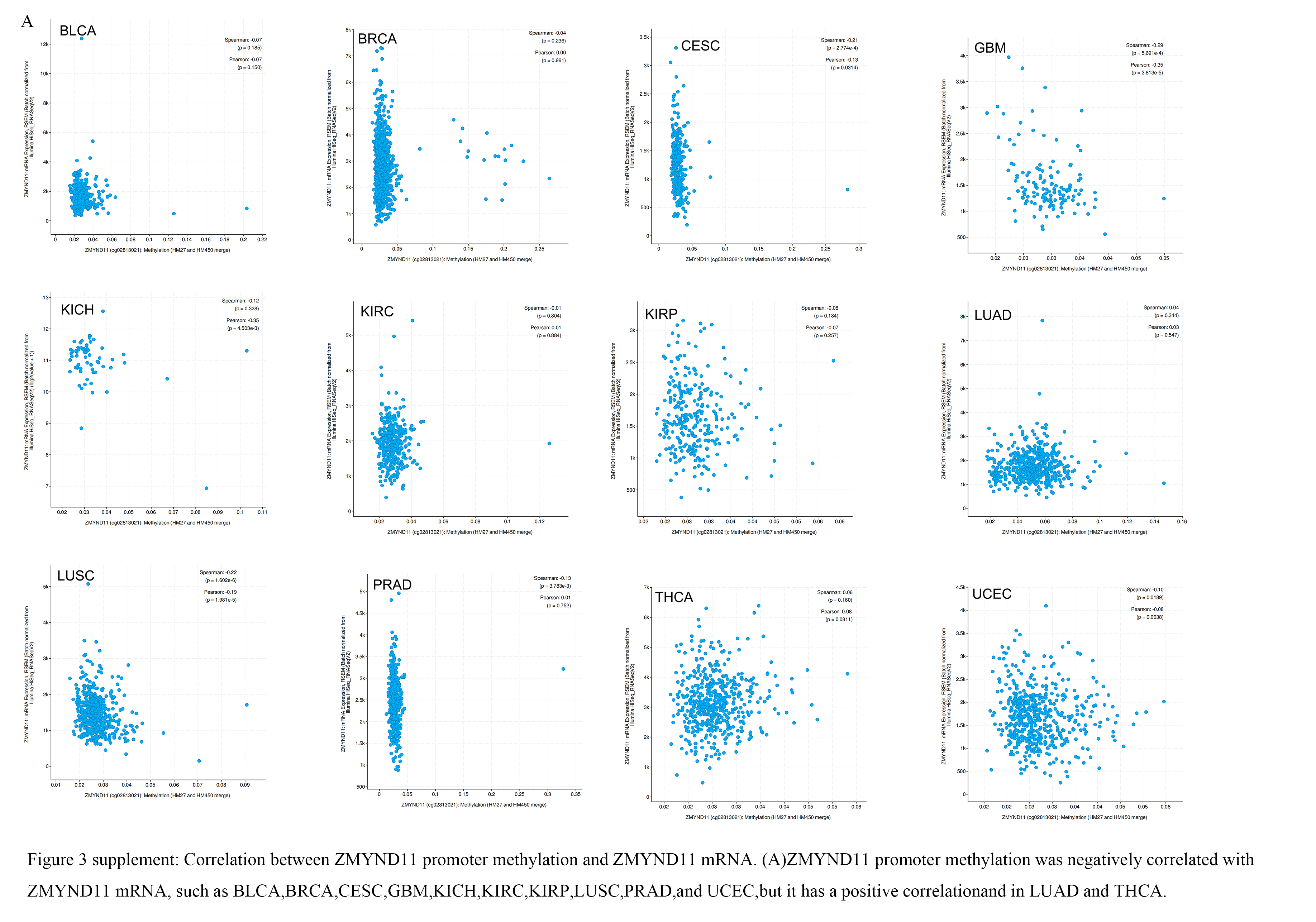
First, we explored the relationship between ZMYND11 methylation in normal and tumor tissues using in TCGA on the ualcan website (Fig. 6A). We subsequently found reduced promoter methylation in the BLCA, BRCA, CHOL, COAD, CESC,ESCA,GBM, HNSC, KIRP, LIHC, LUAD, LUSC, PAAD,PCPG, READ, TGCT,SYAD,THYM,THCA, and UCEC, as well as elevated promoter methylation in the KIRC, PRAD and SRAC. We further analyzed whether ZMYND11 promoter hypomethylation was negatively related to ZMYND11 mRNA expressionin BLCA, BRCA, CESC, GBM, KICH,KIRC,KIRP,LUSC,PRAD and UCEC with low expression in ZMYND11 tumors.But it has a positively weak correlation and in LUAD and THCA(Supplementary Fig. 3A).In LUAD and THCA the low expression of ZMYND11 may be related to promoter methylation.But in other tumors ZMYND11 promoter hypermethylation could not explain the reduced ZMYND11 mRNA, and the low ZMYND11 expression in these tumors may be associated with gene mutations or chromosomal variants that may be associated with other causes.
3.5 Protein phosphorylation analysis data
We demonstrated the comparative discrepancies in the distribution of ZMYND11 phosphorylation levels from normal to primary tumor tissue.The CPTAC set of data revealed that the levels of ZMYND11 phosphorylation in the clear cell renal cell carcinoma, pancreatic adenocarcinoma, head and neck squamous carcinoma, glioblastoma multiforme and breast cancer(Fig. 7A).It demonstrated significant differences in phosphorylation at sites 349, 390, and 393 of ZMYND11 compared to normal tissue (Fig. 7B, p < 0.05). In KIRC at site 349 low phosphorylation, in PAAD at site 390 and 349, in HNSC at site 349 and 390,in GMB at site 349 higher phosphorylation,, in BRCA at site 393 higher phosphorylation.But in PAAD,HNSC and GBM higher levels of phosphorylation sites were exhibited at locus 349.High levels of phosphorylation at locus 349 protein GBM with high total ZNYND11 protein expression.Low levels of phosphorylation at locus 349 protein in KIRC were identified with low total ZNYND11 protein expression.And the functional significance of high levels of ZMYND11 phosphorylation or low levels of phosphorylation in tumour cells may affect the expression of total protein.
3.6 Immuno-infiltration analysis data
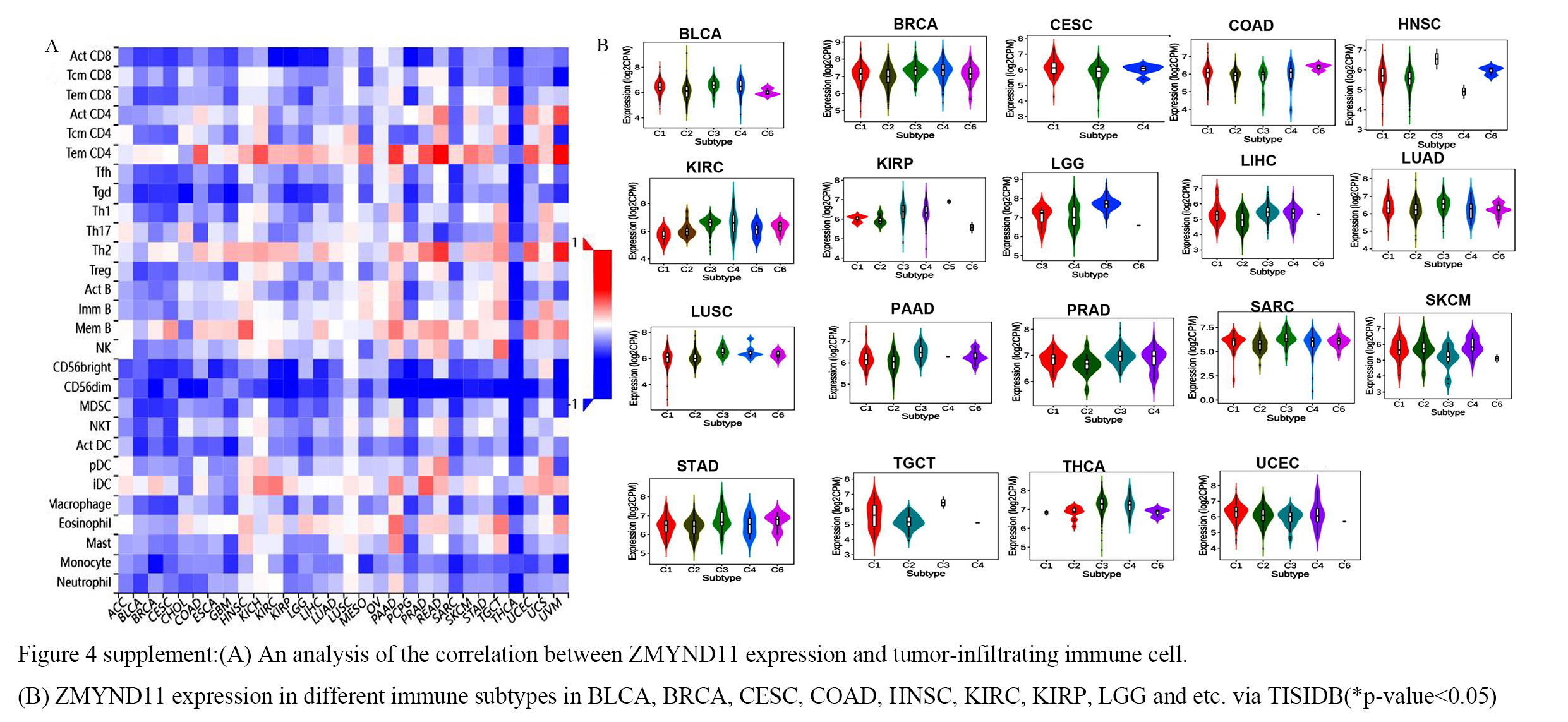
We utilized TIMER2.0 database to have an analysis of the associations between the expression of various immune cells and the ZMYND11 gene in a diverse spectrum of TCGA cancer types based on TIMER, CIBERSORT, CIBERSORT-ABS, QUANTISEQ, XCELL, MCPCOUNTER, and EPIC algorithms. (Fig. 8A and Supplementary table 1), From one series of analyses, according to the EPIC, MCPCOUNTER, XECLL and TIDE algorithms, we determined that the associated fibroblasts had a positive relationship with ZMYND11 expression in the most CESC, COAD, HNSC, HNSC-HPV-, LIHC, LUSC,PAAD, STAD,TGCT and THYM tumors. However ,it negatively correlates with LGG. Subsequently, we applied an algorithm to generate the scatter plot data for the above tumors in the CESC, COAD, HNSC, HNSC-HPV-, LIHC, LUSC,PAAD, STAD, TGCT, THYM and LGG (Fig. 8B), and it was directly related to the infiltration level of cancer-associated fibroblasts based on the one of these algorithms algorithm. According to the a variety of algorithm,We found that the CD8 + T immune cells were positively correlated with ZMYND11 expression in PAAD, STAD andUVM. And neutrophil and endothelial cells were shown to be competitively positively related to ZMYND11 expression in for the majority of tumors(Fig. 9A-C and Supplementary table 1) .
Similarly, we also observed that ZMYND11 expression negatively correlated with ZMYND11 with actCD8 + T, tcmCD8 + T, temCD8 + T, actCD4 + T, and tcmCD4 + T cells. And positively correlated with temCD4 + T cells. This was the same as in the TISIDB database analysis (Supplementary Fig. 4A). Using molecular types of immune subtypes, ZMYND11 mRNA expression was investigated in various immune subtypes.The expression of ZMYND11 mRNA was significantly different in C1 wound healing, C2 interferon-gamma dominant, C3 inflammatory, C4 lymphocyte depleted, C5 immunologically quiet and C6 TGF-β dominant. The differential expression of ZMYND11 in different immune subtypes may partly explain why ZMYND11 shows a variable role in the prognosis of various cancers(Supplementary Fig. 4B).
3.7 Gene enrichment correlation analysis
To determine the role of the ZMYND11 gene in the molecular mechanism of tumorigenesis, we aimed to develop a selection for the targeting of ZMYND11 to the protein, genes interacting with ZMYND11 and related expressed genes, which were subjected to numerous pathways to enrichment analysis. In total, we obtained 50 proteins combined with ZMYND11, as confirmed by experiments using the STRING tool.Fifty proteins associated with the ZMYND11 protein are available in the STRING database. Previous studies have consistently substantiated this finding. As indicated in Fig. 10A, the protein interaction network is displayed. Twenty genes interacting with ZMYND11 were obtained from the GENEMANIA database (Fig. 10B). All available neoplastic analysis data in TCGA were obtained using the GEPIA2 instrument to acquire the preceding 100 genes correlated with ZMYND11 expression. ZMYND11 was positively associated with six genes (KIAA1109,KIF1BP, TTC3, WAC,PURA and FUBP3), as shown in Fig. 10D. The VENN website was used to intersect these three databases to depict common members (Fig. 10C), and it was found that the three databases do not share a common intersection; however, GENEMANIA and STRING have TRAF6, NCOR1, and ETS2, which have been confirmed in the literature. GENEMANIA and GEPIA2 are associated with SOSO2, PJA2, and KIAA1109. These interacting molecules require further investigation.
KEGG enrichment analysis indicated that herpes simplex virus 1 infection, Salmonella infection, and viral carcinogenesis were involved in the ZMYND11 gene effect on tumor mechanisms (Fig. 10E). GO analysis further suggested that genes involved in the ZMYND11 pathway were more involved in RNAase II promoter regulation, positive and negative regulation of the RNAase II promoter, positive and negative regulation of DNA transcription, chromatin modification, and others(Fig. 10F). GO enrichment also indicated that genes interacting with the ZMYND11 pathway were significantly more involved in RNAase II promoter regulation, positive and negative regulation of the RNAase II promoter, positive and negative regulation of DNA transcription, and chromatin modification.
{kind=link}
{kind=link}
{kind=link}
{kind=link}