In geosciences, 3D reconstruction is generally based on the registration of equally-spaced serial sections followed by segmentation of the structures of interest, which is typically done by extracting isosurfaces from the reconstructed image volumes.25 This approach imposes a number of constraints that have to be satisfied in order to get reliable reconstruction results. The first constraint is that registration has to be performed once for all before segmentation. In our experience, however, it is frequent that registration errors manifest themselves only at the visualization stage. Correcting such errors requires a closed-loop pipeline, whereby registration and segmentation can be corrected based on visualization feedback. The second constraint is that the contrast at the boundary of the structures of interest has to be sufficiently large and constant to ensure they can be extracted based on a single intensity threshold. When this not the case, as in the present study, manual contouring followed by contour triangulation may be the only feasible option, which comes at the cost of an increased complexity compared with isosurface extraction when facing complex topological changes. The third constraint is that the serial sectioning has to be performed at a constant spacing. This in turn generally imposes a small spacing to ensure the recovery of details in some regions, even though some regions could be reconstructed using a larger spacing. The number of images to be registered and segmented can become very large. Here, we took advantage of the 3D reconstruction and modelling software Free-D, initially developed for neuroanatomy then cell biology studies26,27 to propose an alternative approach avoiding all these constraints. We show here how using Free-D allowed us to generate insightful 3D reconstructions from manual drawings of sections digitized at arbitrary and varying intervals.
Based on our data, we present evidence and deductions that support the former co-existence of animals that were fossilized together and some speculations on the nature and possible taxonomic relationships of the entombed fossil.
Evidence for the co-existence
Our interpretation of the co-existence of these animals rests on the observations that:
- the entombed shell is situated along the left internal wall of the brachiopod, with its apex close to the umbo of the host; their direction of growth are the same
- maximum length, width and height of the entombed shell is far longer, wider and higher than the maximum gape of the brachiopod valves,
- dimensions of the entombed fossil shell are beyond the maximum gaped width of the brachiopod shells therefore the entombed fossil shell could not have drifted through the commissure of the gaped brachiopod either if it was alive or dead,
- the usually symmetric brachidium of the brachiopod is asymmetric being short on the side of the entombed (=left side),
- the left crus of the brachiopod is shorter, distorted and bent on the side of the entombed fossil
The most convincing evidence for the co-existence of the brachiopod and the captive animal is based on the position, placement and shape of the brachiopod crura. The brachidium is a symmetrically developed structure during the ontogeny.40 All the observed asymmetric deviations of the brachidium may refer to an affection, malformations or deviations caused by internal or external factors.41 Internal (=crural) deformations may appear due to difficult trophic conditions42 that create asymmetry of the brachidium.43 But in most cases both external and internal asymmetry are caused by external factors.44 Internal asymmetric development with symmetric shell also occurs as described at Recent terebratellidinide brachiopods.45 Complete reduction of one lophophore arm is probably connected with directions of food flows43 and are not pathologic. But not in this case. All the observed asymmetric deviations on the crura of the present specimen may refer to a special process that locally and only partially affected the crura.
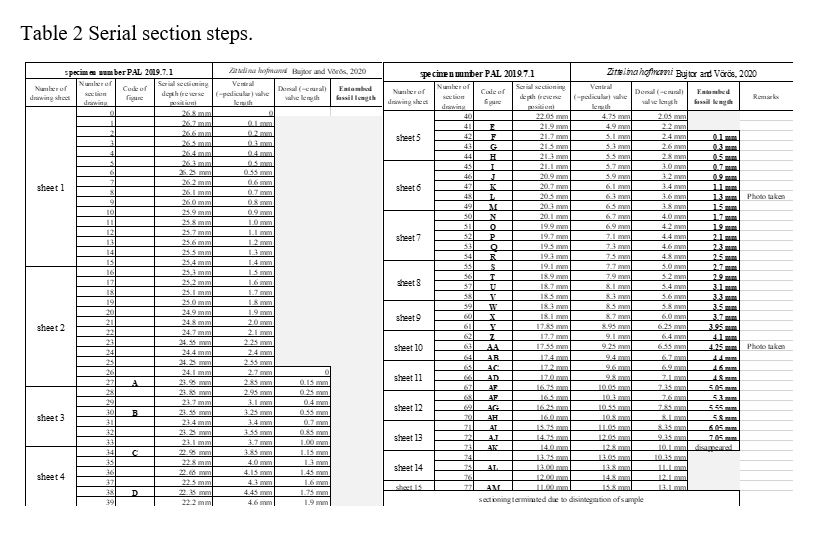
Close to the umbo of the brachiopod (Fig. 4, E: 4.9 to L: 6.3; Fig. 6DE) the left crura is even more developed than its right counterpart, however it is observable (Fig. 4, M: 6.5; Fig. 6AC) that the left brachidium is underdeveloped and shortened. Partial underdevelopment and slight deformation are also present in more posterior regions (Fig. 4, from P: 7.1 to T: 7.9; Fig. 6DE). The most obvious affection on development of the brachidium by the entombed animal is seen on Fig 4, stages from V: 8.3 to Y: 8.95. Here, in the close vicinity of shell of the entombed animal the left brachidium is deformed and bending from the entombed fossil toward the symmetry plane of the brachidium (Fig. 4, V: 8.3) and becomes significantly shorter on the side of the entombed fossil (see on Fig 4, W: 8.5, X: 8.7, and Y: 8.95). These local deformations and asymmetries observed on 2D slices are confirmed by the 3D reconstructions of the whole brachidium, where they appear even more pronounced than in 2D (Fig. 6ACDE). From the point where the width of the entombed shell starts to decrease, the asymmetry of the brachidium of the brachiopod diminishes (as from Fig. 4, AE: 10.05) until the last section with preserved brachidium (Fig. 4, AM: 13.8; see also Fig. 6DE).
The way of entombment
Filter feeding sessile marine animals such as brachiopods, should control incoming water currents in order to filter or prevent any particle or animal to get into their body cavities. This is linked to their anatomy: they need inhalant currents to respirate and filter food particles.46 There are many protective techniques for brachiopods to protect the internal part of their shells.47,48,49,50 Among passive protection devices is the shape of the commissure. Brachiopods and oysters tend to develop zig-zag deflections that function as protective devices51 helping the animal to maintain the same area for influx of sea water as it would without deflections, but significantly decreases the size of grains/eggs able to pass through the commissure.51 In our case, this kingenoid terebratellidine brachiopod did not develop a complicated, zig-zag shaped commissure, therefore any external particle could have penetrated through its commissure more easily. Most probably foraminifers and sponge spicules drifted into these brachiopods via the inhalant current.36 It seems plausible that the animal fossilized inside drifted as an egg into the brachiopod shell.
Other possible ways that the animal got into the shell is by the drifting of a hatched embryo or by the crawling of a juvenile bottom dwelling animal. But brachiopods also have active protective systems as the setae that sense an invader and protect the commissure. This function is well known among living brachiopods,52 however among fossil brachiopods long setae were also present.23,53 It implies that brachiopods may remotely (in a sense of couple of centimetres) control their ambience and easily recognize any invader approaching their commissure early enough to close their valves and prevent the intrusion.
The alternative hypothesis that the fossilized animal specimen was emplaced inside the valves as a corpse seems improbable. An evidence is presented54 that some brachiopods will not open their valves after decay of adductor muscles; however, slightly gaped atrypid brachiopods from the Devonian of Arizona is known.55 Such gaping brachiopods may have provided shelter or food for active benthic organisms.1 In this case it is highly improbable that the shelly animal drifted as a corpse or an abandoned shell into the brachiopod shell because its total length (7.45 mm) and height (3.75 mm) are greater than the slot of the brachiopod’s open commissure. The valves of Recent rhynchonelliform brachiopods stay together after the death of the animal which implies that the entombed fossil was not able to penetrate into the brachiopod even if it was dead. All the above detailed observations show only one plausible solution: the animal that left the fossilized shell inside the brachiopod entered into the body cavity of the living brachiopod as an egg, hatched and developed inside the brachiopod body cavity and left evidence for co-action between the host and the undesired guest.
The possible animal whose shell fossilized inside the brachiopod
This locality that has provided rich and atypically large-sized brachiopods also provided other faunal elements: ammonites, mainly lytoceratids, but some phylloceratids- and haploceratids also occur,8,10 echinoderms,38 crustacean microcoprolites,31,34,36 sponge spicules,10,36 and foraminifers.10,36 Bivalves are missing, and only 3 poorly preserved gastropod remains were collected being rare, but living constituents in this environment; it is plausible that their eggs/larvae may have drifted/crawled into the gaped shells of brachiopods. The average size of encapsulated and shelled veliger gastropod embryos is 742 - 942 μm,56 therefore they could have easily drifted inside gapping brachiopod shells. The size of the hatched crawling gastropod embryos varies between 909 - 1205,56 so they also may crawl inside or have drifted into the brachiopod shell. But crawling into a living brachiopod shell is not probable. Other kinds of gastropod eggs exhibit even smaller dimensions. Many different neogastropod eggs from a deep-sea hydrothermal vent distribute sizes between 100 to 440 μm.57 Although this locality is not a classical deep-sea fossil vent locality but belongs to the rare, shallow marine, sediment hosted hydrothermal vent group (SHV39). A recent analogue to the Zengővárkony fossil environment is provided58 from the submarine hydrothermal vents of the Tyrrhenian Sea (Italy), which suggests that gastropods may have inhabited hydrothermally influenced environments and are the most probable candidates for the entombed fossil. There is another evidence supporting a gastropod: at the apical part of the entombed fossil, a columella-like internal structure is seen on 2D and even 3D reconstructions (Fig. 6). There is evidence for the co-action between the gastropod and the brachiopod. The gastropod influenced the development of the crura of the brachiopod during ontogeny, which became shorter and deviated on the side where the gastropod lived.
{kind=link}