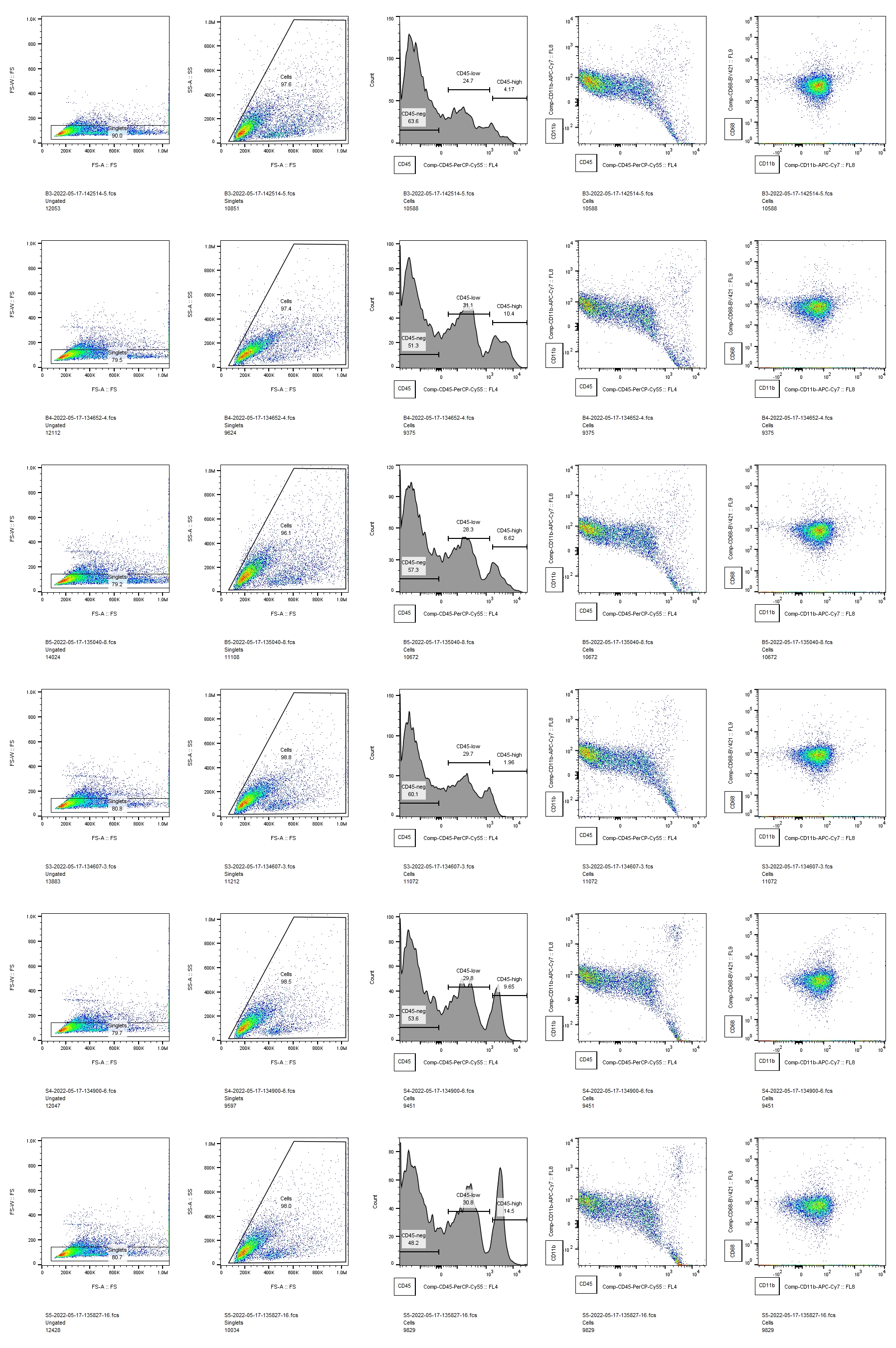
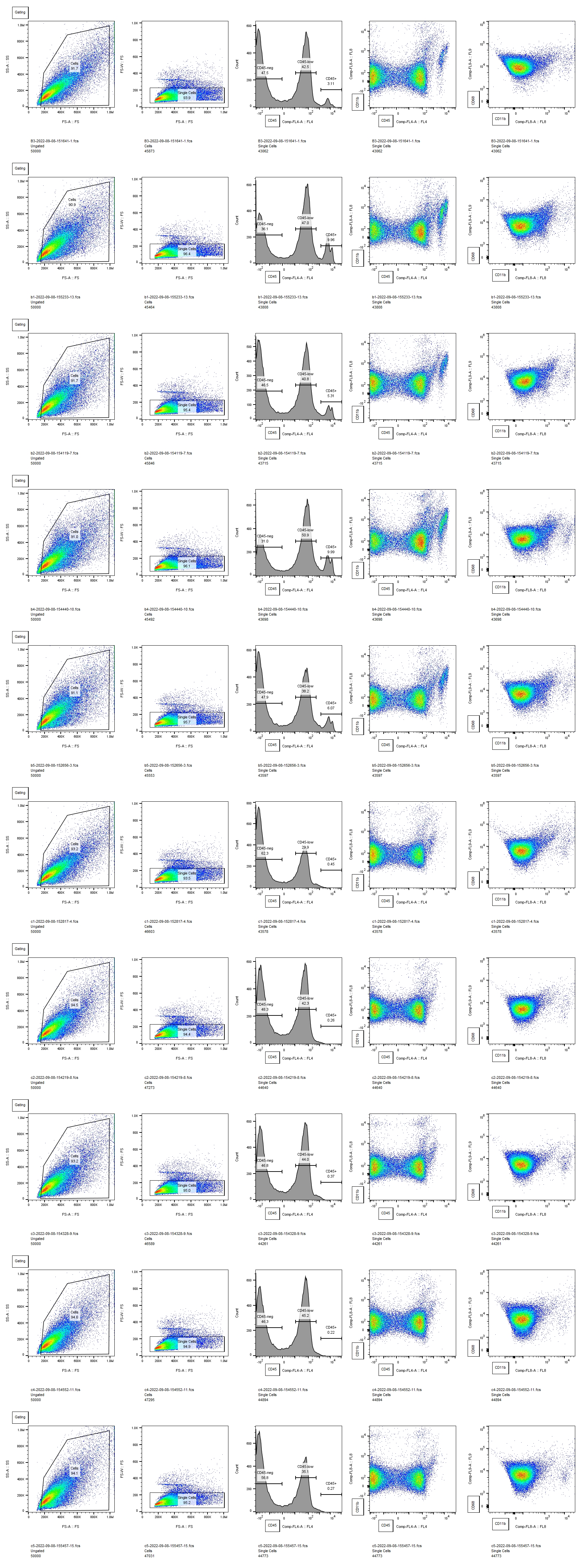
Here, we demonstrated that P5 rat pups mount a more pro-inflammatory innate immune response to IVH and sustain greater white matter injury than P2 pups. This is a particularly robust result, considering that we delivered the same volume of lysed blood into each age animal, resulting in a comparatively larger dose for P2 animals. Our flow cytometry data captures effects throughout the brain, and our previous work has shown that the acute inflammatory response to unilateral IVH is similar on both sides of the brain [3]. In this study, we examined only one time point, but our and other’ prior work showed that inflammation peaks early after IVH when both cytokine analysis and immunohistochemistry are used to quantify the innate immune response [3, 6]. This is similar to a large body of work from stroke and neurotrauma, where the acute inflammatory response peaks early after injury [32, 33]. The markedly different innate immune responses in P2 and P5 animals could be from either endogenous microglia, infiltrating macrophages, or both. However, these experiments were not designed to specifically test whether one type of innate immune cell was the largest contributor to this response. Delineating the differences between endogenous microglia and infiltrating macrophages in the context of development and injury is not straightforward. Antibodies that selectively label microglia may not be reliable in the early developmental period when these experiments were conducted [34] and may be altered by pathology [35, 36]. Furthermore, transient peripheral macrophage infiltration into the brain occurs under normal conditions during the first week of life [37]. Future work will attempt to better distinguish the relative contributions of microglia and peripheral macrophages to the different inflammatory responses detected in P2 and P5 animals. However, we postulate that this is the overall balance of pro-inflammatory cytokines and physiological growth factors that determines white matter fate after IVH rather than the particular cellular source of those factors.
Although white matter injury can be mediated by innate immune cells [8], developing oligodendrocytes are susceptible to direct injury by blood breakdown products [24]. The earlier the developmental stage of the oligodendrocyte, the more susceptible it is to injury from multiple insults [9, 38, 39]. Therefore, our initial hypothesis was that P2 animals would sustain worse white matter injury than P5 animals would. However, we observed more severe white matter pathology in the P5 animals. This suggests that the developmental stage of the innate immune system may be a more critical factor in determining the final white matter pathology after IVH than the developmental stage of white matter. The entire neonatal immune system differs from that of the adult, with an overall dearth of adaptive immune mechanisms compared to mature organisms [40]. Neonates who are born prematurely undergo the transition from an in utero immune system to a more mature immune system in an artificial environment, subject to physiological stressors that can lead to IVH. Clinical experience indicates that hydrocephalus and white matter injury after IVH evolve slowly over time [41]. Microscopically, events that lead to white matter injury are multifactorial and complex. Many molecules present in blood can be toxic to white matter, both directly by interacting with the cell membranes and receptors on OPCs, and indirectly by activating immune cells [1]. The source of these molecules is the intraventricular blood clot that is deposited at the margin of the ventricle at the ictus of IVH. Serial cranial ultrasound studies that are performed routinely in patients with IVH show that these clots gradually lyse and resolve. Therefore, there is likely to be a slow release of injurious factors over time in human IVH. While it is intuitive that the cumulative effects of blood products over time contribute to the ultimate pathology of IVH, our results suggest that the gradual maturation of the immune system over time may also contribute to IVH pathology. If this is the case, a better understanding of how the immune system operates at different ages could help direct surgical and pharmacologic therapy for IVH.
All of our measurements of white matter integrity showed that IVH at P5 induced white matter injury except for the corpus callosum MBP density. Interestingly, our prior work has shown that immunohistochemistry for MBP in the corpus callosum, may not be as sensitive as other measures of white matter integrity such as Western blot.[16] There may also be confounding factors that affect white matter analysis in the corpus callosum but not more peripheral white matter. It is possible that the corpus callosum was depleted of myelin, but due to compression from enlarged ventricles, a reduced amount of MBP was concentrated in a smaller cross-sectional area, making our density measurement less precise. Or the converse maybe true – that the corpus callosum was not significantly depleted of myelin (therefore no change in density), but was stretched as the ventricle enlarged (therefore reduction in corpus callosum thickness). Regardless of the one measure of white matter that was unchanged in the P5 IVH group, our results clearly demonstrate that IVH induced at P5 causes white matter injury that does not occur when IVH is induced at P2.
In addition to white matter injury, IVH also induces hydrocephalus. Most neurosurgeons consider the term “hydrocephalus” to signify increased intracranial pressure from pathological accumulation of CSF. Since we did not study CSF outflow, turnover, or intracranial pressure in this study, it is impossible for us to determine whether our rats had hydrocephalus or just increased ventricular size from white matter loss, as both brain volume loss and hydrocephalus can lead to large ventricles. Therefore, we used the terms “ventricular size” or “ventricular width” to describe the anatomic findings in our study. Future work studying CSF flow will be better able to determine to what extent ventricular changes are due to white matter loss versus obstruction of CSF outflow from the ventricular system. The ventricular enlargement observed here appears more pronounced on histological analysis, likely due to artifacts produced by tissue fixation and slide preparation. On MRI, P2 and P5 control animals, and P2 IVH animals have ventricles that are so small that they cannot be reliably measured. P5 animals that undergo IVH have lateral ventricles that would be considered moderately dilated if compared to human neonates with IVH.
Hydrocephalus and white matter pathology overlap, but are not identical, phenomena as others have eloquently stated [2]. Treating hydrocephalus in animal models can restore white matter integrity [42], suggesting that high intracranial pressure from untreated hydrocephalus may further white matter injury that is not directly caused by inflammatory or cytotoxic effects of blood. Untangling the relative contribution of brain tissue loss and altered CSF dynamics is difficult in experimental IVH models, and even more so clinically [43]. Our results suggest that; the developing neuroimmune milieu may even help to alter the relative contribution of one phenomenon over the course of the disease in individual patients. Improved therapy for IVH will be driven by a better understanding of the pathophysiology of the disease but will also be depend on the acceptance that many complex multisystem diseases are successfully treated via immunomodulatory therapy even before their pathophysiology is completely understood [44].
{kind=link}
{kind=link}