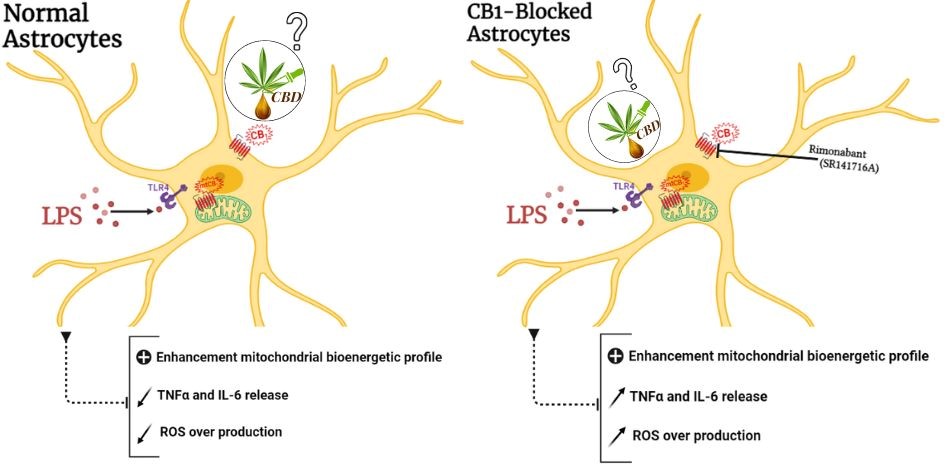
This study aimed to determine (1) how LPS affects the mitochondrial metabolism of astrocytes, and (2) CBD's effect on metabolic activity and pro-inflammatory cytokine release in LPS-treated astrocytes. Our findings show that CBD can modulate the bioenergetic profile of primary astrocytes previously stimulated by LPS, which is an inflammatory-inducing bacterial membrane protein. The CBD-induced bioenergetic modulation consisted of modifying both; oxidative phosphorylation parameters such as mitochondrial membrane proton leak, and mitochondrial spare respiratory capacity, and glycolytic parameters such as glycolytic capacity, and astrocytic glycolytic reserve. The metabolic change induced by CBD was correlated to changes in pro-inflammatory cytokines TNF and IL-6 concentrations. CB1 receptor blocker SR141716A did not impair the anti-inflammatory effect of CBD suggesting that the CBD anti-inflammatory effects may not be mediated by CB1 receptors. Our findings have also shown that CBD reduces ROS production in astrocytes stimulated by LPS. Moreover, SR141716A did not attenuate CBD’s effect on ROS production, suggesting that CBD-induced ROS reduction in LPS-stimulated astrocytes may not be directly mediated by CB1 receptors. Furthermore, our results suggest that CB1 receptors are implicated in the anti-inflammatory response and the regulation of ROS production by LPS-stimulated astrocytes; as CB1 blockade by SR141716A exacerbated the inflammatory response in LPS-stimulated astrocytes by increasing TNF and IL6 levels, as well as ROS production.
LPS-stimulated astrocytes demonstrated a significant decrease in basal mitochondrial respiration that reflects the proportion of OCR used by cells to meet their endogenous ATP demand (Divakaruni et al. 2014). This decrease could be related to a change in the electron chain transport (ECT) activity, in particular, in ECT complex I, the site of NADH dehydrogenase and source of reactive nitrogen (RNS) and reactive oxygen species (ROS) release. These oxidative forms are produced after TLR4 pattern recognition receptor activation by LPS (Lopez-Fabuel et al., 2016) and the activation of their underlying pathways such as mitogen-activated protein kinase (MAPK), myeloid differentiation protein 88-nuclear factor-κB (Myd88-NFκB), and Janus kinases/signal transducers and activators of transcription (JAK1/STAT1) (Gorina et al., 2011). Consequently, triggering the generation of ROS and RNS from NADPH oxidase (NOX) and nitric oxide synthase (iNOS), respectively (Takahashi, 2021). Both ROS and RNS inhibit ECT and decrease mitochondrial membrane potential (Uchi et al, 2014). Recently, it was reported that LPS-stimulated astrocytes activate TLR4 and induce significant production of ROS without RNS production. In astrocytes, the stimulation of TLR4 by LPS induces the activation of the MAPK phosphorylating pathway, underlying ROS release, and subsequent enhancement of PPP flux, as has been suggested by Liyzumi et al., and Takahashi (Iizumi et al., 2016, Takahashi, 2021).
On another hand, we suggest that CBD metabolic effects seem to depend on the inhibition of CB1 receptors by SR141716A, where it significantly decreases proton leak; increases spare respiratory capacity and enhances coupling efficiency in LPS-stimulated astrocytes. In this respect, it was reported that SR141716A significantly decreases CB1 receptor expression and promotes peroxisome proliferator-activated receptor-γ (PPARγ) activation which has a key role in astrocytic metabolism by upregulating glucose, glutamate uptake, and lactate production and release (Wang et al., 2015, Iglesias et al., 2017). These findings support the role of PPARγ activation through CB1 receptors inhibition in CBD anti-inflammatory effects. Moreover, our data indicate that CBD at 1µM prevents LPS-induced mitochondrial damage only when administered in combination with SR141716A. knowing that SR141716A alone does not affect astrocytic metabolism suggests that CBD-induced metabolic effect could be potentiated by negative allosteric modulation of the CB1 receptors as has been suggested by Laprairie et al., (2015). In line with our findings, Sun et al., (2017) confirmed CBD’s effect on mitochondrial bioenergetics in HT22 cells (normal hippocampal neurons) demonstrating that CBD at a dose of 5 µM enhances basal respiration, ATP production-linked oxygen consumption, maximal respiration, and the spare respiratory capacity (Sun et al., 2017).
Lastly, we investigated CB1 receptor inhibition on astrocytic glucose metabolism by measuring the glycolytic flux. Similar to respiratory mitochondrial data, the glycolytic analysis revealed that the glycolytic capacity and glycolytic reserve were increased only following co-administration of CBD with SR141716A while CBD alone had no effect.
Taken together, our data suggest that astrocyte co-treatment with CBD and RS may provide a significant enhancement to both glycolytic capacity and glycolytic reserve under inflammatory conditions induced by LPS-stimulation. It has been shown that CBD effect on astroglial CB1 receptor located on plasma membrane may prevent mitochondrial CB1 receptors (mtCB1R) activation needed for controlling lactate release, and its subsequent related neuronal bioenergetic stress (Jimenez-Blasco et al., 2020). It is well-known that glucose conversion to lactate is critical in the maintenance of neuronal homeostasis (Staricha et al., 2020). Thus, under non-inflammatory conditions astrocytes rely more on glycolysis than oxidative phosphorylation to generate energy and to produce lactate needed by neurons through a mechanism that implicates the endocannabinoid system. Several studies show that CBD has a potent role in mitochondrial chain respiratory regulation and glucose metabolism by enhancing glucose consumption and lactate release (Singh et al., 2015, Sun et al., 2017). However, our investigation outcomes suggest that CBD markedly enhances mitochondrial bioenergetics and promotes protectiveness against LPS-induced inflammation only when applied in combination with CB1 receptor inhibitors.
Besides astrocytes major role in maintaining neuronal energy balance, astrocytes are involved in sustained inflammation in response to infections or diseases (Khandelwal et al., 2011, Klegeris, 2021). It has been shown that astrocytic stimulation by LPS induces a significant increase in proinflammatory markers such as TNF and IL-6 and increases cells' oxidative phosphorylation without affecting their glycolytic metabolism (Krasowska-Zoladek et al., 2007, Jiang et al., 2009, Zamanian et al., 2012). Therefore, we investigated if CBD treatment may prevent the release of these cytokines TNF and IL-6. Our obtained results showed that CBD at a dose of 1 µM significantly decreases TNF and IL-6 release following astrocytic LPS-stimulation (0.2 µM). This result partially corroborates the findings of a recent study showing that CBD indeed attenuates IL-6 release in astrocytes without affecting TNF release (Wu et al., 2021). This discrepancy might be explained, first, by differences in the experimental approaches, as in our study, astrocytes were incubated with LPS (0.2 µg/ml) overnight however in their protocol, astrocytes were stimulated with LPS (0.1 µg/ml) only for 2 h. Secondly, CBD was applied at a dose of 1 µM, however, in Wu et al., study, CBD was applied at a dose of 5 µM. Knowing that CBD has biphasic proprieties, may also explain its lack of effect on TNF release (Wu et al., 2021). Moreover, this difference could be related to the developmental state of the cells (Mecha et al., 2012). CBD's effect on LPS-induced TNF and IL-6 release were not conditioned by CB1 receptor inhibition, by virtue of its effect on metabolism. Thus, CBD’s effect on IL6 and TNF release following LPS stimulation could be mediated by other cannabinoidergic pathways. This hypothesis is supported by other studies showing that CBD's anti-inflammatory effect may involve neither CB1 nor CB2 receptors, In contrast, CBD may act through other pathways such as Wnt3 and NF-κB to inhibit astrocytic cytokine release (Kozela et al., 2010, Esposito et al., 2011, Vallée et al., 2017, Cardinal von Widdern et al., 2020, Wu et al., 2021). However, more studies are needed to confirm these hypotheses. On another hand, regarding CBD well known neuroprotective properties associated with its potent antioxidant capacity (Kim et al., 2021b), our results showed that CBD has the potency to reduce LPS-induced over-production of intracellular ROS in primary astrocytes. This effect is not mediated directly by CB1 receptors activation, as CB1 blockade by SR141716A not counteracted CBD’s effect on ROS release. These results were in line with previous studies demonstrating a similar effect of CBD in LPS-induced oxidative stress in glial cell models. Sonego et al. observed that CBD (10 µM) attenuates intracellular ROS release in primary murine microglia activated by LPS (10 ng/mL) (Sonego et al., 2018). These studies also suggested that CBD-induced attenuation of ROS release might be mediated by its inhibitory effect on the NF-κB signaling pathway. It attenuates NF-κB phosphorylating transcription factor subunit p65 in LPS-stimulated glial cells (Cardinal von Widdern et al., 2020). Also, CBD may inhibit ROS release in LPS-stimulated cells by limiting NADPH production and glucose uptake regarding the fact that glucose is an essential element in the regeneration of NADPH used by NADPH oxidase (NOX) to catalyze ROS production (Kim et al., 2021a, Kim et al., 2021b). Yet, the exact mechanisms by which CBD inhibits the over-production of intercellular ROS induced by LPS remain unresolved.
Furthermore, our study revealed that the application of SR141716A exacerbates TNF and IL-6 release and ROS production by LPS-stimulated astrocytes. This result was in line with findings from a recent study investigating the role of CB1 receptors on the regulation of microglia/macrophage polarization (Lou et al., 2018). This study revealed that the inhibition of the CB1 receptors by SR141716A up-regulates pro-inflammatory cytokines, including TNF and IL-6, and increases nitric oxide (NO) release by microglia/macrophages. Also, this study suggested that the SR141716A effect is related to the TLR-4 and NF-κB/p65 overexpression on microglia/macrophages (Lou et al., 2018). Thus, a next study on the evaluation of NFκB expression in astrocytes could indicate a possible mechanism for CB1.
Our study has provided a new finding alongside the growing evidence that confirms the role of the cannabinoidergic system in the regulation of astrocytes-mediated neuronal circuits homeostasis maintaining, and metabolic support (Covelo A et al., 2021; Iannotti FA et al., 2016; Navarrete M and Araque AJN, 2010). Two lines of data supporting this, first, CB1 receptors expressed in the astrocytic membrane are potential modulators of neurotransmitter reuptake mechanisms by affecting both neurotransmitters and gliotransmitters release (Djeungoue-Petga MA and Hebert‐Chatelain EJB, 2017; Kovács A et al., 2017; Martinez Ramirez CE et al., 2022). Secondly, CB1 receptors expressed in the mitochondrial membrane of astrocytes have a noticeable effect on mitochondrial performance by decreasing ROS release (Achicallende Urcaregui S, 2021; Robledo‐Menendez A et al., 2022). Hence, that opens up novel strategies for exploring interventional therapy for neurodegenerative disorders through modulating astrocytes functions.
Among limitations of our study, is that we don’t explore yet the underlining mechanism. Thus, a study on CB1 knockout mice by using a transgenic model is needed to confirm our hypotheses and greatly extend our knowledge of the molecular mechanisms underlying CBD effects much beyond what has been determined from the CB1 receptor-blocked model by SR141716A. In the current study, CBD effects were evaluated on metabolic and inflammatory parameters in LPS-stimulated primary astrocytes. Therefore, our findings highlight that LPS induced pro-inflammatory cytokines release is mediated by CB1, and this is the crucial point of our study. In this respect, we will conduct a future study to perform more specific experiments related to astrocytic inflammatory response to show how endocannabinoid system could regulate this response.
{kind=link}