GX_P2V(short_3UTR) viral entry through clathrin-coated vesicles (CCV)
To analyze the early interactions between the virus and the host cells at 10 and 30 min post-infection, we used a high multiplicity of infection (MOI) of 10 for this experiment. At 10 min post-infection, we observed a small number of viral particles adhering to the cell surface (Fig. 1B and 1C). These viral particles have a diameter ranging from 50–70 nm and were surrounded by "club-shaped" spikes (Figs. 1B and 1C). The black spots on the virus-like particles are clear (Fig. 1). At 30 min post-infection, GX_P2V was transported into the cell via clathrin-coated vesicles (CCVs) (Fig. 1A). The clathrin-coated scaffold imparts a characteristic bristle coat, which can sometimes be confused with coronavirus-like particles (Additional file 1: Figure S1).
No Significant Differences Observed In Infected Cells At 12 Hours Post-infection
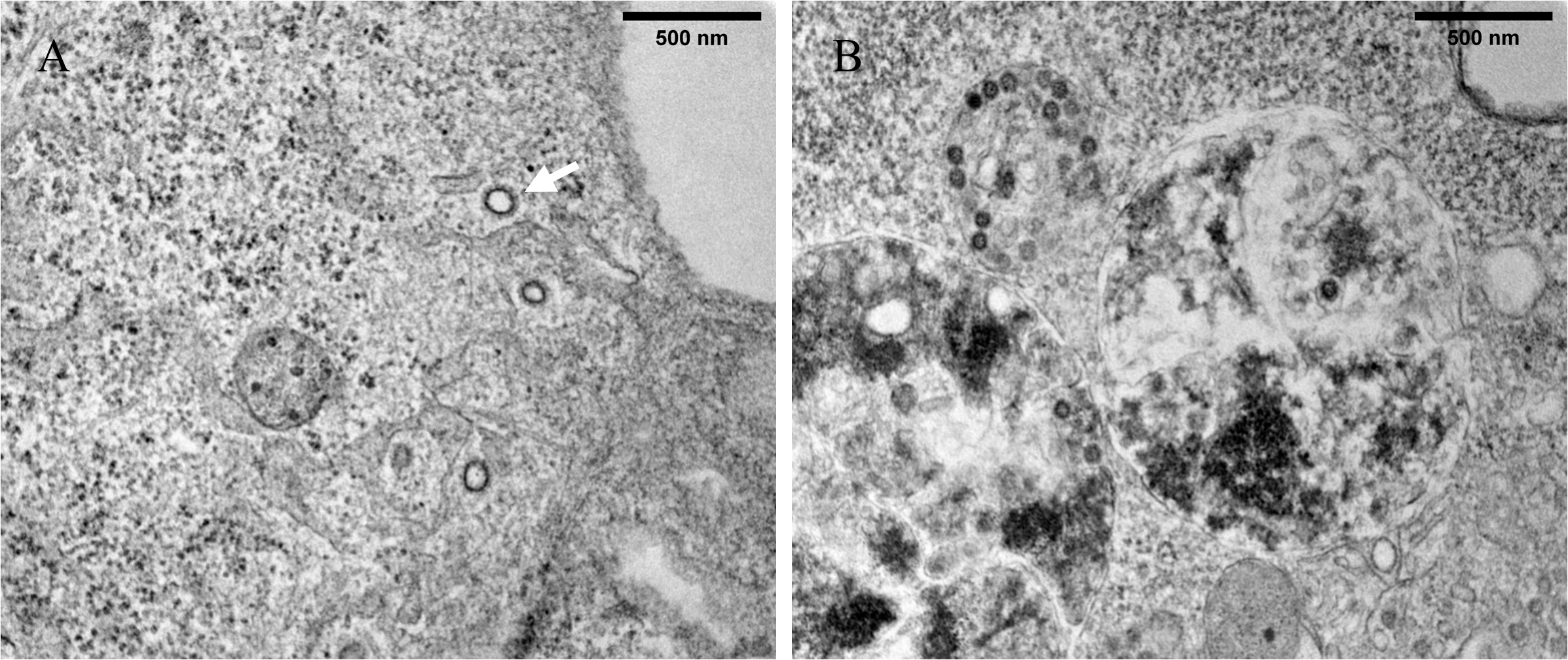
We used a multiplicity of infection (MOI) of 0.1 to analyze the ultrastructural modifications induced by the virus after 12 h post-infection. At 12 h post-infection, we observed that GX_P2V(short_3UTR)-infected cells were not substantially different from uninfected cells, and no obvious alterations in cellular structures were seen (Fig. 2). This is in stark contrast to SARS-CoV-2, which can form a large number of DMVs in cells before 12 h post-infection[14, 15]. This suggests that the replication speed of GX_P2V(short_3UTR) might be much slower than that of SARS-CoV-2. The delayed formation of numerous vROs might be one of the mechanisms contributing to the attenuation of GX_P2V(short_3UTR).
Viral Replication Organelle Structures In Gx_p2v(Short_3utr)-infected Cells At 24 Hours Post-infection
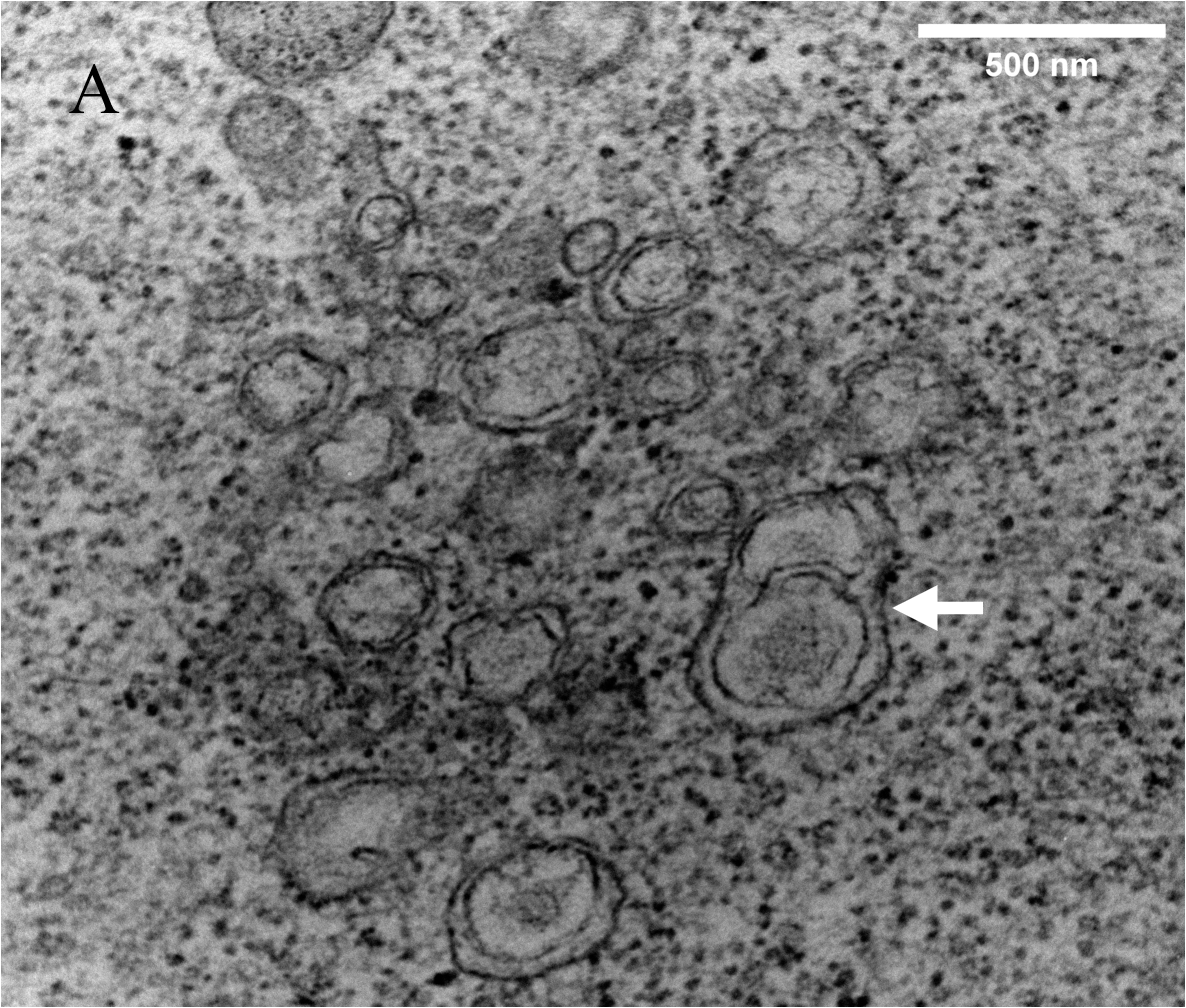
At 24 h post-infection, we observed significant ultrastructural changes in GX_P2V(short_3UTR)-infected cells, including the formation of DMVs, DMSs, and CMs, which collectively comprise the viral replication organelle (vRO) (Fig. 3). DMVs varied in size (100–350 nm in diameter) and were characterized by a double membrane and distinct outer membranes (Fig. 4A). The outer membranes of two DMVs fused with each other (Additional file 1: Figure S2). DMSs were more homogeneous in size (around 70 nm in diameter) and resembled β-CoVs structures (Fig. 4A). Both DMVs and DMSs were surrounded by mitochondria and the latter exhibited high electron density.
CMs appeared as winding, maze-like structures derived from the ER (Fig. 4A). We also observed annular lamellae (AL) consisting of highly ordered stacks of ER-derived membranes containing nuclear pore complexes (NPCs) arranged in parallel (Fig. 4B). NPCs, initially believed to be characteristic of rapidly dividing cells, have also been identified in + RNA virus-infected cells[16].
Viral particles with prominent spikes were observed budding in ERGIC (Fig. 5). And particles in large intracellular vacuoles (Fig. 6) were potentially associated with the autophagy-lysosome pathway. The number of viral particles in GX_P2V(short_3UTR)-infected cells was significantly lower than that observed in SARS-CoV-2-infected cells reported by others[14]. Additionally, we observed one cell compartment with extended microtubules (Fig. 6B).
Late-stage Infection: Reticulovesicular Network And Lysosomal Release Of Gx_p2v(Short_3utr)
At 48 h post-infection, GX_P2V(short_3UTR) induced notable cytopathic and cellular ultrastructural changes, with abundant GX_P2V(short_3UTR)-induced membrane modifications present in the cells (Fig. 7). A portion of the cytoplasm appeared to be disintegrating (Fig. 7A), and numerous cell vacuoles containing few or no virus-like particles were observed (Fig. 7B).
Compared to 24 h post-infection, the virus induced the formation of an elaborate reticulovesicular network (RVN) (Fig. 8), with the structure peaking at 48 h post-infection. DMVs were not observed at this stage, but numerous vesicles with diameters ranging from 100 to 300 nm were encapsulated in large vesicle packets (VP) (Fig. 8A).
Many virus-like particles accumulated in multivesicular bodies (MVBs), with diameters ranging from 0.4 to 1 µm (Fig. 9). Endosomal or lysosomal-sized single membrane vesicles were observed fusing with MVBs containing the virions (Fig. 9A).
Like other β-CoVs[9], GX_P2V(short_3UTR) appears to utilize the lysosomal pathway for export. TEM images revealed GX_P2V(short_3UTR) accumulation in vesicles released by exocytosis (Fig. 10). The infected cell seems to strip a piece of cytoplasm (Fig. 10A), and multiple autophagy-like vesicles were present within the cells (Fig. 11A).
Released GX_P2V(short_3UTR) particles were present on the cell surface (Fig. 11) and between cell surfaces (Fig. 11B), displaying distinct crowns or spikes. Importantly, the number of viral particles in all the observed vacuoles was low, consistent with observations at 24 h post-infection, and in contrast to the high numbers observed in SARS-CoV-2-infected cells[14]. This finding suggests a potential link to GX_P2V(short_3UTR) attenuation and highlights the differences in replication and release between GX_P2V(short_3UTR) and SARS-CoV-2.
{kind=link}
{kind=link}